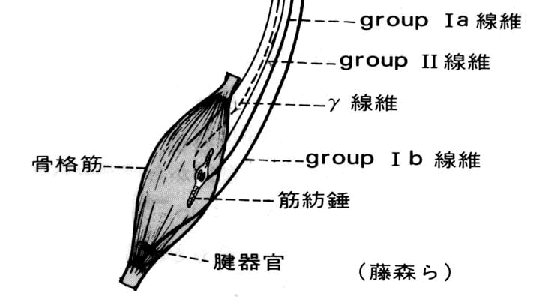
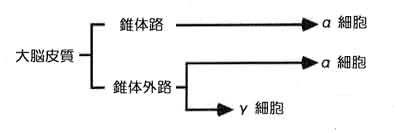
戞侾恾乮恾夝惗棟妛丒堛妛彂堾傛傝乯
嬝朼悗偺旈枾
嬝擏憖廲朄偺扙椡偺巇曽傊偺曉帠
僯僼僥傿偺僷僜僐儞捠怣偺拞偵丄娍曽偺晹壆偲偄偆僼僅乕儔儉偑偁傝傑偡丅偦偺拞偺乽娍曽偺廃曈乿偲偄偆夛媍幒偑偁傝丄巹偼偦偙偵搳峞偟偰偄傑偟偨丅偙偺俫俹偺乽偁傞偠偺晹壆乿偺暥復偼丄戝晹暘偑丄偦偙偱巹偑搳峞偟偨婰帠偑傕偲偵側偭偰偄傑偡丅
偦偺夛媍幒偱goopon偝傫偲偄偆摨偠嬒惍巘偺曽偲乽嬝擏憖廲朄偺扙椡偺巇曽乿偵偮偄偰乮#964,966,971乯堄尒傪岎姺偟傑偟偨丅岎姺偲偄偭偰傕丄斵偺偄偭偰偄傞棟榑偑惓偟偄偺偐娫堘偭偰偄傞偺偐丄偝偭傁傝傢偐傜側偄偺偱丄岎姺偵傕側傝傑偣傫丅
傑傞偱丄挻尰幚庡媊偺奊傪尒偰偄傞傛偆偱偡丅崅偄寶暔偐傜悈偑壓偵棳傟偰偄偰丄偦偺棳傟傪偳傫偳傫偨偳偭偰峴偔偲丄壓偵壓偭偰偄偨悈偑丄偄偮偺娫偵偐寶暔偺忋偺尦偺応強偵栠偭偰偄傞丄偲偄偆奊偱偡丅妋偐偵悈偼壓偵棳傟偰偄傞偺偵丄偨偳偭偰峴偔偲丄偄偮偺娫偵偐忋偵側偭偰偄傞丅偳偙偱娫堘偭偨偺偐暘偐傜側偄丅
偙偺奊偼丄偙傫側偙偲偼尰幚偵偁傝摼側偄偺偱丄柧傜偐偵偙偪傜偑嶖妎偟偰偄傞偲暘偐傞偺偱偡偑丄goopon偝傫偺庡挘偼尰幚偲堦抳偟傑偡丅壗偲側偔娫堘偭偰偄傞傛偆側婥偑偡傞偺偱偡偑丄斵偺庡挘傪偨偳偭偰峴偔偲丄傗偭傁傝惓偟偄偺偐側丄偲巚偭偰偟傑偄傑偡丅斀榑偟傛偆偵傕嬝擏偵偮偄偰偺怺偄堛妛揑抦幆偑側偄巹偵偼丄擄峌晄棊偺忛偺傛偆偵寶偪傆偝偑偭偰偄偰斀榑傪備傞偟偰偼偔傟傑偣傫丅
揇撽幃偵嬝擏偺廃曈偺抦幆傪曌嫮偟偰傕丄堛妛梡岅偑擄夝偱丄偝傜偵偙偺斀幩偵娭偡傞晹暘偼丄偄傠偄傠側奣擮偑傑傞偱栐偺栚偺傛偆偵娭楢偟偰偄傞偺偱丄撉傒恑傫偱偄傞偆偪偵丄摢偺拞偑偛偪傖偛偪傖偵側傝丄壗偑壗偩偐暘偐傜側偔側偭偰偟傑偄傑偟偨丅
偦偺傛偆側傢偗偱丄夞摎傪墑婜偟偰偄傞偆偪偵丄憗侾擭敿偺嵨寧偑偨偭偰偟傑偄傑偟偰丅堦帪偼傗傔偰偟傑偍偆偐丄偲巚偭偨偺偱偡偑丄捈娤偱偼壗偐戝帠側傕偺偑偙偙偵愽傫偱偄傞婥偑偟偰偄偨偺偱丄嬒惍朄抋惗50廃擭廽偆徏嶳偱偺慡崙島廗夛偵娫偵崌傢偣傞傋偔丄傕偆堦搙僠儍儗儞僕偟偰傒傑偟偨丅偙傟偼侾擭敿墇偟偺仈971偺僐儊儞僩偵懳偡傞夞摎偱偡丅
杮棃偼僯僼僥傿偺曽偵搳峞偡傋偒傕偺偱偡偑丄僷僜僐儞捠怣偱偼夋憸偑巊偊傑偣傫丅崱夞偺夞摎偵偼懡検偵夋憸傪巊傢側偗傟偽側傜側偄偺偱丄巹偺俫俹傪巊傢偞傞傪摼傑偣傫偱偟偨丅偛椆夝偔偩偝偄丅
971/971 VYL05551 Goopon RE:嬝擏憖廲朄偺扙椡偺巇曽丂(10) 99/10/30 04:06 965傊偺僐儊儞僩
價乕僶乕偝傫丅偙傫偵偪偼丅埲壓丄挿暥偱怽偟傢偗偁傝傑偣傫丅嬝擏憖廲朄偵偮偄偰丄傑偨峫偊偰傒傑偟偨丅偲傝偁偊偢偺寢榑偲偟偰偼丗
侾丏懠摦揑偵備偭偔傝堷偒怢偽偟丄媧婥偺捀揰偱懴偊傜傟傞偩偗懴偊偝偣偰乽僷僢偲扙椡乿偟偨曽偑丄乽備偭偔傝扙椡乿偡傞傛傝傕抩娚嶌梡偑嫮偄
俀丏侾偺憖朄傛傝傕摍広惈廂弅偺曽偑抩娚嶌梡偑嫮偄
俁丏俀偺憖朄傛傝傕摍挘惈廂弅偺曽偑抩娚嶌梡偑嫮偄
偲尵偊偦偆偱偡丅扐偟丄忬嫷偵傛偭偰嵟揔側憖朄偼堎側傝傑偡丅侾偼摉弶偺媈栤傪書偄偨偲偒偲偼斀懳偱嬒惍朄偺憖朄捠傝偵側偭偰偄傑偡丅
夁嫀偺敪尵偲廳暋偟傑偡偑丄傕偆堦搙婎杮揑側抦幆傪惍棟偝偣偰偄偨偩偒傑偡丅
崪奿嬝偺嬝慄堐偼悗奜嬝慄堐偲悗撪嬝慄堐偵暘偗傜傟傑偡丅
壗偩丄側傫偩丄摍広惈廂弅丄摍挘惈廂弅丄悕奜嬝丄悕撪嬝偰丅
摍広惈廂弅
帺傜廂弅偡傞嬝擏偵懳偟丄懠摦揑偵掞峈傪壛偊偰偦偺嬝挿傪曄壔偝偣側偄傛偆偵偟偨廂弅
嬝偺椉抂傪屌掕偟偰巋寖偡傞偲丄廂弅拞嬝偺挿偝偼曄壔偟側偄丄偦偺傛偆側廂弅丅嬝偺堦抂偵挘椡婰榐憰抲傪偮偗傟偽丄挘椡曄壔傪婰榐偡傞偙偲偑偱偒傞丅偡側傢偪摍広惈廂弅偱偼抁弅嬋慄偼摼傜傟偢丄挘椡嬋慄偑摼傜傟傞丅乮堛妛戝帠揟乯
憡庤偑弅傔傛偆偲偡傞偺偵丄偙偭偪偑掞峈傪偔傢偊偰弅傔側偄傛偆偵偝偣傞傢偗偩丅憡庤偲偍側偠椡偑昁梫偩偹丅憡庤偑偳傫偳傫椡傪弌偟偰弅傔傟偽丄偙偭偪傕椡傪弌偡偐傜丄偦偺椡傪寁傟偽偳傫偳傫忋偑偭偰偄偔傢偗偐丅偦傟傪挘椡嬋慄偲偄偆偺偩偹丅傛偆偡傞偵丄嬝擏偺挿偝傪曄偊側偄傛偆偵椡傪壛偊偰廂弅偝偣傞傢偗偩丅榬憡杘偟偨傜椉幰偼摨偠埵抲偱椡傪壛偊偰峴偔偺偩偹丅傑偨偼丄峧堷偒偱榬傪怢偽偟偨傑傑峧傪堷偔忬懺丠丅
摍挘惈廂弅
摍挘惈廂弅丗帺傜廂弅偡傞嬝擏偵懳偟丄偦偺廂弅椡傛傝庛偄掞峈椡傪懠摦揑偵壛偊偨忬懺偱嬝挿偑抁偔側傞廂弅
嬝偺堦抂傪屌掕偟丄懠抂偵堦掕偺廳傝乮晧壸乯傪偐偗偰巋寖偡傞偲丄廂弅拞嬝偼晧壸傪堷偒忋偘傞偙偲偵傛偭偰廂弅偡傞丅摍挘椡偲偄偆偺偼廂弅拞嬝偺挘椡偑晧壸偵摍偟偄堦掕偺抣偵側偭偰偄傞偐傜偱偁傞丅乮堛妛戝帠揟乯
嬝擏傪弅傔傞椡傪堦掕偵偟偰挿偝傪曄偊傞傢偗偩偹丅榬憡杘偱偼晧偗偰偄傞忬懺丠丅
悗撪嬝慄堐
兞塣摦偺慄堐偺妶摦偵傛偭偰廂弅偡傞嬝慄堐偱丄嬝朼悗乮嬝怢挘庴梕婍乯偺拞偵懚嵼偟丄廂弅妶摦偵傛傝丄嬝朼悗偺姶搙傪挷愡偡傞栶栚傪傕偮丅( 堛妛戝帠揟乯
悗奜嬝慄堐
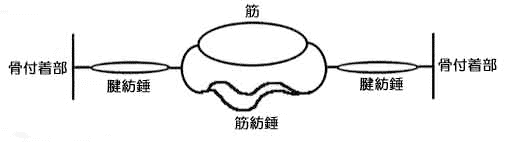
堦曽丄兛塣摦恄宱偱巟攝偝傟偰偄傞嬝慄堐乮戝晹暘乯傪悗奜嬝慄堐丂extrafusal f侾倐er丂偲偄偆丅乮恾夝惗棟妛乯
側乕傫偩丄悕奜嬝偲偄偆偺偼偨偩偺嬝擏丄悕撪嬝偲偄偆偺偼嬝朼悗偺偙偲偐丅
悕奜嬝丂丂偨偩偺嬝擏乮偄偄壛尭偩偹乕乯
悕撪嬝丂丂嬝朼悗乮嬝擏偺拞偵偁傞挘椡専弌婍丄僙儞僒乕偺堦庬偩偹乯
偦傟偵偟偰傕丄堛妛梡岅偼擄偟偄丅堛幰偲偄偆偺偼堦斒恖偵傢偐傜側偄尵梩傪巊偭偰乽偍傑偊偨偪傛傝巹偨偪偺曽偑執偄傫偩偲乿桪墇姶偵怹偭偰偄傞恖庬偠傖側偐傠偆偐丅偦傟偵偙傫側偵尵梩偑擄偟偄偲巊偭偰偄傞杮恖偨偪傕傢偐傜側偔側傞傫偠傖側偄偐側丅傑乕偄偄偐丅
悗奜嬝慄堐偼崪奿偵晅拝偟偰偄傞偺偱廂弅偡傞偲椡傪敪惗偟傑偡丅偙偺悗奜嬝慄堐偺椉抂偵偼捈楍偵銯婍姱乮銯朼悗乯偑埵抲偟偰偄傑偡丅
嬝擏偺抂偺銯偺堦晹偵偼銯朼悗偲偄偆丄偙傟傕嬝擏偺嬞挘搙傪儌僯僞乕偡傞僙儞僒乕偑偁傞丄偲偄偆偙偲偹丄偙傟偼椆夝丅
堦曽丄悗撪嬝慄堐偼悗奜嬝慄堐偲暲峴偟丄崪奿嬝偺姶妎婍偲偟偰嶌梡偟偰偄傑偡偑丄椉抂偑幚幙揑偵偼帺桼抂側偺偱廂弅偟偰傕椡傪敪惗偟傑偣傫丅
側傫偱傛丅偙傟偵偼恾偑昁梫偩偹丅
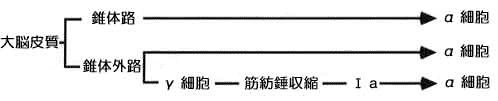
戞侾恾乮恾夝惗棟妛丒堛妛彂堾傛傝乯
側傞傎偳帺桼抂偲偄偆偺偼丄晜偄偰偄傞丄偲偄偆偙偲偐丅崪偵偮偄偰側偄偐傜椡傪敪婗偱偒側偄傢偗偹丅偦偟偰丄嬝擏偺挘椡乮挘傝嬶崌乯偺寁應憰抲偩偐傜丄嬝擏杮棃偺摥偒偼偟側偄偲偄偆偙偲偩側丅
嬝擏偺廂弅偲抩娚偵娭學偡傞恄宱偵偮偄偰偼丄戙昞揑側傕偺偲偟偰媮怱惈偺侾倎偲侾倐丄墦怱惈偺兞偲兛偵拲栚偟傑偡丅埲壓丄乽悗撪乿偲乽悗奜乿偵拲堄偟偰撉傒恑傔偰偔偩偝偄丅乮恄宱傪侾倎丄侾倐偲昞婰偟傑偟偨偑丄捠忢丄侾偺晹暘偼儘乕儅悢帤偺厽偱彂偒昞偝傟傑偡丅偟偐偟丄偙傟傜偼婡庬埶懚暥帤偺偨傔丄Mac仺Win偱偼乽厽厾叀參乿偵丄Win仺Mac偱偼乽嘥嘦嘨嘩乿偵側偭偰偟傑偄傑偡偺偱丄偁偊偰嫟捠偺侾乣係偵曄偊傑偟偨丅婥偵側傞傛偆偱偟偨傜丄僟僂儞儘乕僪屻丄専嶕仺抲姺偊偺婡擻偱偁傜偐偠傔曄姺偟偨曽偑椙偄偱偟傚偆丅乯
偙傟偼暥復偠傖傢偐傝偵偔偄偐傜丄偙傟偵傕恾偑昁梫偩側丅
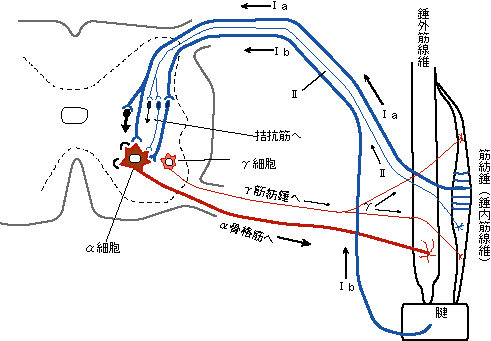 丂
丂
戞俀恾
側傞傎偳偹丅偙偆側偭偰偄傞偺偐丅彮偟婥偵側傞偺偼嬝朼悗偺壓偺抂偑銯偵偔偭偮偄偰偄傞偲偙傠偩偹丅戞侾恾偱偼嬝朼悗偼銯偵偼偮偄偰偄側偄丅偙傟偼嶲峫偵偟偨恾偑偁傑傝偵傕彫偝偄偺偱丄巹偑僀儔僗僩儗乕僞偱戝偒偔彂偒捈偟偨傕偺偩丅偙偺尦恾偼僐僺乕偺僐僺乕丄偦偺傑偨懛偺懛丄偁偨傝偐傜偲偭偰偄傞偺偱丄尦偼堛妛彂側偺偩偑斉尦偑傢偐傜側偄丅偱傕丄堛妛彂偱偙偆側偭偰偄傞偺偩偐傜丄娫堘偭偰側偄偲巚偆丅偦傟偵丄偙偺晹暘偼偁傑傝廳梫偱偼側偄偲巚偆丅
兛偼悗奜嬝慄堐傊廂弅乛抩娚柦椷傪揱払偟傑偡丅
兛嵶朎偐傜偼兛恄宱慄堐偑弌偰偄傞偑丄偙傟偑嬝擏偵廂弅偺柦椷傪弌偡偺偩側丅偩偗偳抩娚柦椷偲偄偆偺偼側傫偩丅嬝擏偼廂弅偟偐偟側偄偱偼側偄偐丅偮傑傝帺傜偼怢傃側偄丅偡傞偲丄抩娚柦椷偲偄偆偺偼丄怣崋偑棳傟側偄丄偡側傢偪侽偺忬懺側偺偩傠偆丅偦傟偼兛嵶朎偑嫽暠偟偰偄側偄偲偄偆偙偲偠傖側偄偺偐側丅
偱偼丄廂弅偟偨嬝擏偼偳偆偟偨傜怢傃傟傞偺丅偦傟偵偼漢峈嬝偑廂弅偟偰丄懳峈偡傞嬝擏傪怢偽偟偰偄傞偺偩傠偆丅偰側傢偗偱丄偙偙偱偼丄抩娚柦椷偲偼丄怣崋偑棳傟側偄丄擖椡乽侽乿偲偟偰棟夝偟偰偍偙偆丅
侾a偼悗撪嬝慄堐偺拞墰偵廔枛偟偰偄偰嬝擏偑怢傃偰偄偔帪偺乽挿偝偺曄壔乿乮悗奜嬝慇堐偲悗撪慇堐偺挿偝偺嵎乯偲偦偺乽懍搙乿偵斀墳偟丄挿偝偑抁偔側傞帪偵偼斀墳偟傑偣傫丅
亀乮悗奜嬝慇堐偲悗撪慇堐偺挿偝偺嵎乯亁偰側傫偩丅嬝擏偺挿偝偲嬝朼悗偺挿偝偺嵎丠丄偙傫側偺斾妑偟偰偳傫側堄枴偑偁傞偺偩傠偆丅尵偄偨偐偭偨偙偲偺堄枴偼丄師偺傛偆側偙偲偐側丅
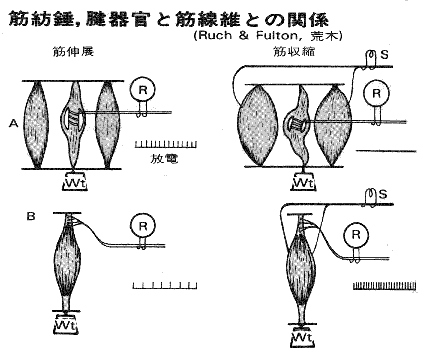
戞俁恾乮恾夝惗棟妛乯
戞俁恾忋抜偱偼嬝朼悗偼嬝擏偑廂弅偡傞偲丄嬝朼悗傕弅傫偱偟傑偭偰丄挿偝傪姶偠傜傟側偔側偭偰偄傞乮忋抜塃偱偼曻揹傪傗傔偰偟傑偭偰偄傞乯丅嵍偱偼嬝擏偑怢傃傞偲曻揹傪偟偰傞丅偙偺偙偲傪偄偭偰偄傞偺偐側丅傕偭偲傕兞偑偐傜傫偱偔傞偲丄偝傜偵嬝朼悗傕弅傓偺偩偗傟偳丄偙傟傪偄偭偰偄傞偲傗傗偭偙偟偔側傞偺偱屻夞偟乮屻夞偟1乯丅偱傕丄乽悗奜嬝慇堐偲悗撪慇堐偺挿偝偺嵎乿傪姶偠傞偵偼偦偆偟側偄偲偱偒側偄偺偩偐傜丄偨傇傫偦偺偙偲傪偄偭偰偄傞偲巚偆偺偩偗傟偳丅
侾倎偺斀墳偼捈愙偦偺嬝擏偺兛偵嫽暠柦椷傪弌偡偺偱丄偦偺嬝擏傪廂弅偝偣傛偆偲偟傑偡丅
傕偆彮偟徻偟偔偄偆偲丄侾倎偵揹棳偑棳傟傞偲丄兛嵶朎偑嫽暠偟丄偦偺寢壥兛慄堐偵怣崋偑棳傟丄偦偺怣崋偑嬝擏傪廂弅偝偣傞丄偲偄偆偙偲偩側丅
侾倐偼銯婍姱偵廔枛偟偰偄偰乽嬝挘椡乿偵斀墳偟丄愐悜恄宱傪夘偟偰偦偺嬝擏偺兛偵梷惂柦椷傪弌偡偺偱丄偦偺嬝擏傪抩娚偝偣傛偆偲偟傑偡丅
彮偟栠偭偰丄戞俁恾偺壓抜傪傒偰傒傞偲壓抜偼銯朼悗偺恾偩丅嬝擏偼僂僄僀僩傪偐偗傜傟偰堷偭挘傜傟偰偄傞偑丄堷偭挘傜傟怢傃偰偄傞偲偒偱傕曻揹偟偰乮嵍乯丄嬝擏偵柦椷偑偒偰乮揹棳傪棳偟偰傞乯嬞挘偟偰偄傞帪偵傕曻揹偟偰偄傞丅乽乽嬝挘椡乿偵斀墳偟乿偲偄偆偺偼丄偙偺傛偆側偙偲偐側丅偱傕側丄嬝偺挘椡偵斀墳偡傞偲偄偆偙偲偼丄侾倎乮嬝朼悗乯偩偭偰峀偔峫偊傟偽嬝偺挘椡偵斀墳偟偰偄傞偺偩偗傟偳偹丅挘椡傊偺斀墳偺巇曽偑堘偆偺偩傠偆側丅銯朼悗偺曽偑嫮偄挘椡偵側傞偲斀墳傪偼偠傔傞偲偄偆偺偑惓妋偱偼側偄偐側丅
偩偑丄栤戣偼
亀愐悜恄宱傪夘偟偰亁偰偳偆偄偆偙偲偐側丅偩偭偰丄侾倎傕侾倐傕傒傫側愐悜偵擖偭偰傞傛丅偦偺拞偵偁傞兛嵶朎偵偵宷偑傞偨傔偵擖偭偔傞偺偩偐傜丄傢偞傢偞侾倐偺傒亀愐悜恄宱傪夘偟偰亁偲丄偙偲傢傞昁梫偼側偄偱偼側偄偐側乕丅
扨側傞尵梩偺偁傗側偺偐側丅憡斀恄宱斀幩偲崿摨偟偰偄側偄偐偺偐側丅偙傟側傜丄亀愐悜恄宱傪夘偟偰亁偄傞丅偙偺栤戣偼乽廂弅偟偐偟側偄嬝擏偑偳偆偟偨傜怢傃傟傞偺乿偲偄偆媈栤偵傕偵偮側偑傞偹丅

戞係恾乮恾夝惗棟妛乯
俙丗巐摢嬝乮俻乯偺嬝朼悗乮俙俽乯偐傜偺侾倎慄堐偑丄俴俇惛悜儗儀儖偵偍偄偰丄巐摢嬝塣摦僯儏乕儘儞偵扨僔僫僾僗惈偵楢棈偟丄偦偺懁巿偑偍側偠愐悜晹偵懚嵼偡傞侾倎梷惂惈夘嵼僯儏乕儘儞偵廔傢傝丄偙偺夘嵼僯儏乕儘儞偺幉嶕偑俴俈儗儀儖偺擇摢嬝斀銯條嬝乮俛俽俿乯偺塣摦僯儏乕儘儞偵懳偟偰梷惂嶌梡傪媦傏偡宱楬傪帵偡丅
戞係恾偼憡娭恄宱巟攝傪偟傔偟偰偄傞丅偙傟偼旼奧銯斀幩偺椺偩偗傟偳丄旼偺偍嶮偺壓偺戝戁巐摢嬝偺銯傪僩儞僇僠偱寉偔懪偮偲丄懌偑挼偹忋偑傞丅偦偺儊僇僯僘儉偺愢柧丅偨偨偐傟偰戝戁巐摢嬝偑廂弅偡傞偲丄摨帪偵斀懳懁偺戝戁擇摢嬝偑娚傑側偗傟偽側傜側偄丅偦偆偟側偗傟偽儘僢僋偝傟偨忬懺偵側偭偰偟傑偭偰丄懌偑偼偹偁偑傞偙偲偼側偄丅偙偺傛偆側庡摦嬝偱偁傞戝戁巐摢嬝偲漢峈嬝偱偁傞戝戁擇摢嬝偺娭學偺恾偩傛丅偙偺娭學偑憡娭恄宱巟攝偲偐憡斀恄宱斀幩偲偄傢傟傞傕偺偩丅
偙傟傪恄宱偺棳傟偱尒傞偲丄戝戁巐摢嬝偺廂弅偵傛傝丄偦偺怣崋偑侾a恄宱傛傝俴俇愐悜偵揱傢傝乮媮怱乯丄俴俇愐悜偱偼偦偺怣崋傪庴偗偰丄堦晹傪堦偮壓偺俴俈愐悜偵憲傝丄偦偺怣崋偑俴俈愐悜偺兛慄堐乮墦怱乯偵揱払偝傟丄偦傟偑戝戁擇摢嬝偵揱偊傜傟傞偙偲偵傛偭偰擇摢嬝偑備傞傓丅偙偺椺偩偲丄怣崋偑亀愐悜恄宱傪夘偟偰亁偲偄偆偺傕棟夝偱偒傞丅
偮偄偱偵偄偭偪傖偆偲丄嬝擏偑怢傃傞偺偼偙偺巇慻傒側偺偱偡丅斀懳偺嬝擏偑弅傫偱丄尦偺嬝擏偑怢傃傞偺偱偡丅嬝擏偼帺傜偼廂弅偟偐偟側偄偺偱偡丅
偱傕丄偙傟偼侾倎偱婲偙傞偙偲側偺偱丄侾倐偱偼婲偙傜側偄丅偙偙偱偼偦偆偄偆姩堘偄偱偼側偄偲偟偰丄愭偵恑傕偆丅偙傟偼幹懌偩偭偨偹丅
兛偵梷惂柦椷傪弌偡偺偱丄偦偺嬝擏傪抩娚偝偣傛偆偲偟傑偡丅
兛偵梷惂柦椷傪偳偆傗偭偰弌偡偺偐側丅偩偭偰侾倐偵怣崋偑棳傟偰偄傟偽丄偦傟偼兛嵶朎偵撏偒丄偦傟偵傛傝兛嵶朎偑嫽暠偡傞偐傜丄兛慄堐偵傕怣崋偑棳傟丄嬝擏偼嬞挘偡傞偺偱偼側偄偺丅戞俀恾偵傕偳偭偰丄侾倐偺棳傟傪傛偔尒偰傒傞偲丄偁傟丄侾倐偲兛嵶朎偺娫偵壗偐嫴傑偭偰傞偧丅偙傟側傫偩丅
偦偆偐丄偙偆偄偆偺傪夘嵼僯儏乕儘儞偲偄偆偺偐丅夘嵼僯儏乕儘儞偺拞偵偼梷惂惈僯儏乕儘儞偲偄偆偺偑偁傞偺偩側丅梷惂惈僯儏乕儘儞偺傎偐偵偼嫽暠惈僯儏乕儘儞偲偄偆傕偺傕偁傞偺偐丅側傞傎偳偹乕丅
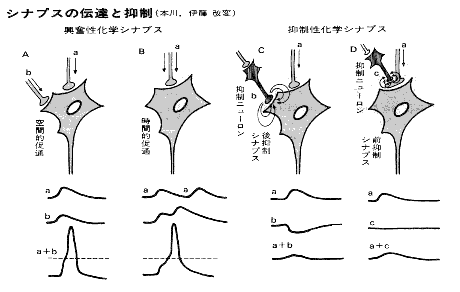
戞俆恾乮恾夝惗棟妛傛傝乯
梷惂惈僔僫僾僗乮堛妛戝帠揟丒堛帟栻弌斉乮姅乯乯
僔僫僾僗慜偺嫽暠偑揱払偝傟傞偲僔僫僾僗屻僯儏乕儘儞偺嫽暠揱払傪偍偝偊傞傛偆偵嶌梡偡傞僔僫僾僗傪偄偆丅僔僫僾僗屻梷惂偲僔僫僾僗慜梷惂偺擇偮偺宆偑偁傞丅慜幰偼僔僫僾僗屻枌偵捈愙偵梷惂揱払暔幙偑嶌梡偟丄梷惂惈僔僫僾僗屻揹埵傪惗偠偰僔僫僾僗屻僯儏乕儘儞偺妶摦揹埵偺敪惗傪梷偊傞傕偺偱偁傞丅
屻幰偼丄梷惂惈僯儏乕儘儞偑嫽暠惈僯儏乕儘儞偺幉嶕枛抂晹偵僔僫僾僗傪嶌偭偰偄偰丄慜幰偺妶摦偺寢壥丄嫽暠惈僯儏乕儘儞偺妶摦帪偵偍偗傞幉嶕枛抂偐傜偺揱払暔幙偺曻弌偑尭彮丄偦偺寢壥丄嫽暠揱摫偑梷惂偝傟傞傕偺傪偄偆丅
偱丄梷惂偺寢壥偼偳偆側傞偺偩丅恄宱嵶朎偺拞偵擇偮偺巋寖偑擖偭偰偐傜丄倎偺巋寖偲倐偺巋寖偑偍屳偄偵憡嶦偟崌偆偺偑屻梷惂丄恄宱嵶朎偵擖傞捈慜偱僇僢僩偟偰偟傑偆偺偑慜梷惂偐丅偙偺偄偢傟偐偱侾倎偼梷惂偝傟偰偄傞偺偩偹丅偨傇傫嵍偐傜俁斣栚偺僔僫僾僗屻梷惂偺曽偩傠偆側丅
侾倐偺斀墳偼擻摦揑廂弅帪偵憹壛偟傑偡丅侾倎偼侾倐傛傝鑷抣偑掅偄偺偱嫽暠偟傗偡偄偱偡丅
偙偺擻摦揑偲偄偆尵梩丄偄偮暦偄偰傕側偠傑側偄側丅側傫偩偭偗丅偦偆偐庴摦偺斀懳偩偐傜乽帺暘偐傜摥偒偐偗傞偙偲乿偐丅偦偆偡傞偲丄侾倐偼嬝擏偑帺暘偐傜弅傫偩帪偵斀墳傪憹壛偝偣傞偲尵偆偙偲偩側丅帺暘偱弅傫偩偲偒偵偼堦斣嫮偄揹棳傪棳偡偲偄偆偙偲偩側丅偦傟偼慜偺恾偺壓抜嵍偱傕柧傜偐偩偹丅偱傕丄偦傟偩偗偠傖側偄偧丅嫮偔堷偒怢偽偟偰傕斀墳偡傞偧丅偱傕丄擻摦偺斀懳乽庴摦乿偱偼偳偆側傞偺偐側丅
偁傟丄庴摦偲偄偆偙偲偼乽懠恖偵弅傔傜傟偨傝丄堷偒怢偽偝傟傞乿偲偄偆偙偲偩傛側丅偦偆偟偨応崌偼丄斀墳偑庛偔側傞偺偐丅偦傫側偙偲柍偄偺偱偼丅銯朼悗偼僙儞僒乕偩傛丅僙儞僒乕偑懠恖偐帺暘偐幆暿偱偒傞偺偐偹丅尵梩偵堷偭偐偐偭偰傞偲愭偵恑傑側偄偗偳丄堦墳僠僃僢僋丅
戞俇恾乮恾夝惗棟妛乯
僑儖僕銯婍姱偲偄偆偺偼銯朼悗偺暿柤丅戞俇恾傪傛偔尒傞偲丄嬝擏偑捠忢忬懺偱偼銯朼悗偼敪幩偟偰側偄丅怢傃傞偐丄弅傓偐偟偨偲偒偩偗敪幩偟偰偄傞偹丅偦偟偰丄嬝朼悗偼怢傃偰偄傞偲偒偵敪幩偟偰丄嬝擏偑弅傫偩偲偒偵偼敪幩偟偰偄側偄丅偦偟偰丄偨傇傫擻摦傕庴摦偱傕丄偡側傢偪帺暘偱弅傕偆偑怢傃傛偆偑丄懠恖偑弅傑偣傛偆偑怢偽偦偆偑丄敪幩乮敪壩偲傕偄傢傟偰偄傞偑乯偵偼娭學側偄偲巹偼巚偆丅
侾倎偼侾倐傛傝鑷抣偑掅偄丅嬝朼悗偺曽偑晀姶偵斀墳偡傞偲偄偆偙偲偩偹丅梫偡傞偵嬞挘姶偵偼嬝朼悗偺曽偑傛傝姶偠傗偡偔偰丄銯朼悗偼撦偄偲偄偆偙偲偩側丅
偦傝傖偦偆偩丄偨傇傫侾倐偑斀墳偡傞偲偄偆偺偼丄媫寖偵堷偒怢偽偝傟偨傝偟偰嬝擏偵尷奅埲忋偺椡偑壛傢偭偨偲偒丄偮傑傝梊婜偟側偄偲偒偵揮傫偩傝偟偨偲偒側偳偵斀墳偡傞丅偦偟偰枵懷傗嬝偑愗傟偰偟傑偆偺傪杊偖偨傔偵偁傞偺偱偼側偄偐側丅偄傢偽嬝擏偺埨慡憰抲偩傛丅偩偐傜丄捠忢偺忬懺偱偼侾倎偑摥偄偰丄旝柇側嬝偺僐儞僩儘乕儖傪偟偰偄傞偺偩偐傜丅侾倎偺鑷抣偑崅偗傟偽偦傟偼偱偒側偄偐傜偹丅
兞偼嬝擏偑廂弅偟偨帪偵悗撪嬝慄堐偑偨傞傫偱悗奜嬝慇堐偲偺挿偝偺嵎偑専弌偱偒側偔側傝丄嬝朼悗偺姶搙偑掅壓偡傞偙偲偑側偄傛偆偵悗撪嬝慄堐傪廂弅偝偣傑偡偑丄椡偼敪惗偟傑偣傫丅
妋偐偵丄嬝擏傪弅傔傞椡偼側偄偑丄偙傟廳梫偩傛丅偙傟傪柍帇偟偪傖偄偗側偄丅偟偐偟丄愭偵恑傒偨偄偺偱丄偙傟傕屻夞偟乮1乯偲偮側偑傞偹丅
兛偵偼侾倎偐傜偺廂弅(+)柦椷偲侾倐偐傜偺抩娚(-)柦椷偑摨帪偵揱傢傝丄偦偺弖娫弖娫偵柦椷偺嵎偟堷偒傪偟偰丄偦偺寢壥廂弅柦椷偑桪惃偱偁傟偽廂弅偟丄抩娚柦椷偑桪惃偱偁傟偽抩娚偟傑偡丅偦偺掱搙偼嵎偟堷偒偺寢壥偺愨懳抣偵墳偠傑偡丅
乽侾倐偐傜偺抩娚(-)柦椷乿偲偄偆偺偼丄惓妋偵偼梷惂僯儏乕儘儞傪夘偟偨抩娚偩側丅傕偭偲傕乽嬝擏傛怢傃傠乿偲偄偆柦椷偼側偄偐傜丄偨偩扨偵乽嬞挘偟傠乿偲偄偆柦椷傪丄偍偝偊偰僠儍儔偵偟偰偄傞偵偟偐偡偓側偄偺偩偑偹丅
偟偨偑偭偰丄嬞挘偟偰偄傞嬝擏傪抩娚偝偣傛偆偲偡傞応崌丄側傞傋偔侾倎偺斀墳偑庛偔偰侾倐偺斀墳偑嫮偔側傞傛偆偵嬝擏傪憖嶌偡傞偲岠壥偑戝偒偄偲梊憐偱偒傑偡丅
偙傟偑丄偄傑偄偪傢偐傜傫丅
侾倎偺斀墳偑庛偄丂仺嬝朼悗傪庛偔斀墳偝偣傞丠丂仺嬝傪庛偔堷偭挘傞丠乮斀墳偝偣側偔偡傞偵偼嬝擏傪弅傔傝傖偄偄偺偩偑丠乯
侾倐偺斀墳偑嫮偄丂仺銯朼悗傪嫮偔斀墳偝偣傞丂丂仺嬝擏傪嫮偔堷偭挘傞偐嬞挘偝偣傞丂仺梷惂僯儏乕儘儞偱兛偼巋寖偝傟側偄
嬝朼悗傪斀墳偝偣側偔偡傞偵偼嬝擏傪廂弅偝偣傟偽傛偄丅偦偺廂弅忬懺偱銯朼悗傪斀墳偝偣傞偵偼丄丒丒丒偁乕偩傔偩丄僀儊乕僕偱偒側偄丅傕偆彮偟愭偵恑傔偽尒偊傞偺偐側丅
偙傟傜偺抦幆傪尦偵嬝擏偺斀墳傪扨弮壔偟偰峫偊偰傒傑偡丅
傑偢丄乽懠摦揑偵備偭偔傝嬝擏傪怢偽偟偨屻丄扙椡乿偡傞憖嶌偱偼丄埲壓偺傛偆偵側傝傑偡丅
側偍丄挘椡偑侾倐偺鑷抣埲忋偺応崌丄偙偙偱偼懄侾倐桪惃偲偟偰埖偄丄偙偺寢壥傪挘抩偲偟偰昞帵偟偰偄傑偡丅幚嵺偵偼侾倎偲侾倐偲偱偳偪傜偑桪惃偐敾抐偟側偗傟偽側傝傑偣傫偑丄捀揰傑偱怢挿偡傟偽偳偙偐偱昁偢侾倐偑桪惃偵側傞偲壖掕偟扨弮壔偟偰偁傝傑偡丅
挘抩偵偍偄偰廂弅傪乽亄乿丄抩娚傪乽亅乿偲偟偰崌寁傪嶼弌偟偰偄傑偡丅偨偩偟丄抣偼掕惈揑側孹岦傪偮偐傓偨傔偺栚埨偵夁偓傑偣傫丅
乽懠摦揑怢挿偲扙椡偺応崌乿
憖嶌(1)丗嬒惍朄偺嬝擏憖廲朄偺捠傝丄備偭偔傝偲嬝擏傪怢偽偟偰偄偒丄怢偽偟愗偭偨捀揰偱僷僢偲扙椡偟偨応崌偱偼埲壓偺傛偆偵側傝傑偡丅

側傞傎偳丅摦偐偟側偑傜憖嶌偟偰偄傞傢偗偐丅
憖嶌(2)丗(1)偲摨條偵捀揰傑偱怢挿偟丄搑拞傑偱備偭偔傝扙椡偟丄師偄偱僷僢偲扙椡偟偨応崌

憖嶌(3)丗(1)偲摨條偵捀揰傑偱怢挿偟丄嵟屻傑偱備偭偔傝扙椡偟偨応崌
側傞傎偳偹乕丅傛偔峫偊偨傕傫偩偹丅
扙椡夁掱偵偍偄偰悗撪嬝慇堐偵兞偐傜廂弅柦椷偑揱傢偭偨偲偟偰傕丄悗奜嬝慇堐偵偼捈愙娭學偟側偄偺偱忋婰偺寢壥偵塭嬁偼偁傝傑偣傫丅
偆乕傫丄偙偙偼偳偆偐側丅偙傟傕屻夞偟乮1乯偵偮側偑傞偹丅
偟偨偑偭偰丄憖嶌(2)偲(3)偑(1)傛傝傕抩娚嶌梡偑嫮偦偆偱偡丅偮傑傝丄扙椡夁掱偺侾倐偺鑷抣埲忋偱摼傜傟傞抩娚嶌梡傕棙梡偱偒傞偲偄偆偙偲偱偡丅偙傟偑埲慜偺敪尵偱乽僷僢偲扙椡乿偡傞傛傝傕乽備偭偔傝扙椡乿偟偨曽偑抩娚嶌梡偑嫮偄偺偱偼側偄偐偲峫偊偨棟桼偱偡丅
祩﹤祦A埲忋偺憖嶌偵偼帪娫偺奣擮偑濨枂偱偡丅幚嵺偵偼庴幰偺屇媧傗惍幰偺帩媣椡側偳偵傛傝帪娫揑惂尷偑偁傝傑偡丅戝掞偼庴幰偺屇媧偱惂尷偝傟傞偲巚偄傑偡丅偦偙偱尷傜傟偨帪娫撪偱側傞傋偔侾倎偺斀墳偑庛偔偰侾倐偺斀墳偑嫮偔側傞傛偆偵嬝擏傪憖嶌偡傞偵偼丄偲峫偊傞偲埲壓偺傛偆偵側傝傑偡丅
憖嶌(4)丗(1)偲摨條偵捀揰傑偱怢挿偟丄媧婥偺尷奅傑偱懴偊偰扙椡偟偨応崌

億僀儞僩偼乽媧婥偱懴偊傜傟傞偩偗懴偊偝偣偰乿偵偁傝傑偟偨丅偮傑傝丄乽捀揰乿偡側傢偪乽侾倎偑嵟傕斀墳偣偢丄侾倐偑嵟傕嫮偔斀墳偟偰偄傞忬懺乿傪偱偒傞偩偗挿偔堐帩偝偣傞偙偲偱偡丅憖嶌(4)偱偼揔摉偵捀揰傪俁偮暲傋偰傒傑偟偨丅嬒惍朄偱偼屇媧偑媡偵側傞偲嶌梡傕媡偵側傞偺偱屇婥偵擖傞弖娫偵乽僷僢偲扙椡乿偡傞偺偩偲巚偄傑偡丅
傑偨丄嬝擏偺婲巒晹偲掆巭晹偵庤傪摉偰偰堷偒怢偽偡傛偆偵偡傞偙偲偱丄妋幚偵銯婍姱偵挘椡傪壛偊傞偙偲偑偱偒傑偡丅偙傟偱惗棟妛揑儗儀儖偱偺媈栤偑夝寛偟傑偟偨丅
婥偵側傞偺偼億僀儞僩偺梌偊曽偲崌寁偩偹丅廂弅偲抩娚偵億僀儞僩傪梌偊偰崌寁偱弌偟偰偄傞偗傟偳傕丄偼偨偟偰偙傟偑嬝擏偺乽備傞傒乿偵宷偑偭偰備偔偺偐側丅偙傟傕屻偱弎傋傞偙偲偵偟傛偆丅屻夞偟乮2乯
偟偐偟丄侾倎傗侾倐偺斀墳偼惗懱奜偱偺幚尡偱摼傜傟偰偄傞抦幆偺條側偺偱丄幚嵺偺惗懱撪偱憖嶌(4)偺尰徾偑偳偺掱搙婲偒偰偄傞偐偼掕偐偱偼偁傝傑偣傫丅
偦偆側傫偩傛丄彍擼摦暔偺嬝擏傪巊偭偰挘椡偺幚尡偦偟偰偄傞傛偆側偺偩偗傟偳傕丄偙傟偭偰巰懱偺嬝擏偺偙偲偩傠偆丅惗偒偰偄傞偺偲偩偄傇堘偆偺偱偼側偄偐側丅
崱丄尒捈偟偰傒傞偲偙傟傜偺巋寖朄偼奆丄壗傜偐偺宍偱棊嵎傪棙梡偟儊儕僴儕傪偮偗偰偄傑偡丅偦傟偵傛偭偰嶖岆偵傛傞拞棫揰偺僘儗傪儕僙僢僩偟偰弶婜抣偵傕偳偟偰偄傞傛偆偵巚偊傞偺偱偡丅
寢壥揑偵偼尒帠偵價乕僶乕偝傫偺嬄傞偲偍傝偺憖朄偵側偭偰偄傞傛偆偱偡丅
妋偐偵拞棫揰偺儕僙僢僩偲偄偆偙偲傪偄偭偨傛丅偦偺堄枴偼乽杽傕傟偰偄偨庤媄椕朄偺摑堦尨棟俀亅 斀幩曇乮http://www.linkclub.or.jp/~jingyuu/aruji/touitu2.htm乯乿偱弎傋偰傑偡丅
偲偙傠偱丄侾倎偼嬝擏偑怢傃偰偄偔帪偺挿偝偺曄壔偲偦偺懍搙偵斀墳偡傞偲彂偒傑偟偨偑丄幚嵺偵偼挿偝偵曄壔偑側偔偰傕掕忢揑偵庛偔斀墳偟偰偄傑偡丅偙傟傪峫椂偟偰傕忋婰偺寢壥偵杦偳塭嬁偼偁傝傑偣傫丅
偙偺侾倎偺摿挜傪媡庤偵棙梡偡傞偲丄侾倎偼嬝擏傪怢偽偝側偗傟偽斀墳偟偵偔偄偲尵偊傑偡丅偙偺掕忢揑側斀墳傪侾倎'偲昞傢偟丄戝嶨攃偵亄0.2乮偙偺抣偼暥專傛傝侾倎偺扙暘嬌偺昿搙偐傜媮傔傑偟偨乯偲偟丄偦偺嵎偑弌傞偐傕偟傟側偄憖嶌偵偮偄偰師偵峫偊偰傒傑偡丅
摍広惈廂弅丗擻摦揑偵廂弅偡傞嬝擏偵懳偟懠摦揑偵掞峈傪壛偊偰偦偺嬝挿傪曄壔偝偣側偄傛偆偵偟偨廂弅
梫偡傞偵丄嬝擏偺挿偝傪曄偊側偄傛偆偵椡傪壛偊偰廂弅偝偣傞傢偗偩丅庴幰偵帺傜嬝擏傪弅傔偝偣偰丄惍幰偑偦傟偵掞峈傪壛偊傞偲偄偆偙偲偩偹丅乽嬝挿傪曄壔偝偣側偄傛偆偵乿偲偄偆偙偲偼庴幰偺椡偲惍幰偺椡偑摨偠偱側偗傟偽偄偗側偄偲傕偄偊傞偹丅椡斾傋傪僀儊乕僕偡傟偽傛偄偺偐側丅
憖嶌(5)丗摍広惈廂弅傪偝偣捀揰偱懴偊傜傟傞偩偗懴偊偝偣偰丄僷僢偲扙椡偟偨応崌

丂
偟偨偑偭偰丄摍広惈廂弅傪棙梡偟偨応崌乮憖嶌(5)乯偺曽偑懠摦揑怢挿傪棙梡偟偨応崌乮憖嶌乮3乯乯傛傝傕抩娚嶌梡偑嫮偔側傝傑偡丅
峏偵丄侾倎偼擻摦揑偵嬝擏偺挿偝傪弅傔偰偄傞夁掱偱傕兞偵傛傝姶搙掅壓傪曗彏偝傟偰偄傑偡偑丄摍広帪傛傝偼斀墳偟偵偔偔側傞偺偱丄偙偺惈幙傪棙梡偡傞偙偲傪峫偊偰傑偡丅
偙偺帪偺侾倎傪侾倎"丄偦偺抣傪亄0.1偲偟傑偡丅
摍挘惈廂弅丗擻摦揑偵廂弅偡傞嬝擏偵懳偟偦偺廂弅椡傛傝庛偄掞峈椡傪懠摦揑偵壛偊偨忬懺偱嬝挿偑抁偔側傞廂弅
偙傟偼丄嬝擏傪弅傔傞椡傪堦掕偵偟偰挿偝傪曄偊傞傢偗偩偹丅庴幰偑椡傪擖傟偰嬝擏傪弅傔傛偆偲偡傞偺偵懳偟丄偦傟傛傝傕庛偄椡偺掞峈傪壛偊傞丅椡偔傜傋偱偼晧偗偰偄傞偺偱丄嬝偼弅傓丅憖懱朄傪僀儊乕僕偡傟偽椙偄偹丅
憖嶌乮6乯丗摍挘惈廂弅傪偝偣捀揰偱懴偊傜傟傞偩偗懴偊偝偣偰丄僷僢偲扙椡偟偨応崌

偟偨偑偭偰丄摍挘惈廂弅傪棙梡偟偨応崌乮憖嶌乮6乯乯偺曽偑摍広惈廂弅傪棙梡偟偨応崌乮憖嶌(5)乯傛傝傕抩娚嶌梡偑嫮偔側傝傑偡丅偨偩偟丄憖嶌(5)偲乮6乯偱偼侾倐偺抩娚嶌梡偑丄挘椡偺捀揰傑偱偺傕偭偰峴偒曽丄捀揰偺嫮偝丄漢峈偝偣傞掞峈椡偺嫮偝側偳偵傛偭偰旝柇偵曄壔偡傞偺偱堦奣偵尵偊傑偣傫丅
偙偙傑偱嶰捠傝偺嬝擏偺廂弅丒怢挿朄偵偮偄偰尒偰偒傑偟偨偑丄扨弮偵侾倐偺恄宱廔枛傪捈愙埑敆偡傞偙偲偱銯婍姱偵捈愙挘椡傪惗偠偝偣傞偙偲偑偱偒傞偺偱丄侾倐偺斀墳傪婲偙偝偣傞偵偼桳岠偲峫偊傜傟傑偡丅
埲忋偺寢壥偐傜師偺傛偆側偙偲偑尵偊偦偆偱偡丅
抩娚嶌梡偵偮偄偰丗
仠側傞傋偔侾倎偺斀墳偑庛偔偰侾倐偺斀墳偑嫮偔側傞傛偆偵嬝擏傪憖嶌偡傞偲岠壥偑戝偒偄丅偦偺孹岦偲偟偰偼乽銯婍姱偺埑敆亙懠摦揑怢挿亙摍広惈廂弅亙摍挘惈廂弅乿偩偑丄愨懳揑側傕偺偱偼側偄丅
埨慡惈偵偮偄偰丗
仠懠摦揑側怢挿偱偼惍幰偑揔愗側挘椡傪姶抦偱偒側偗傟偽丄庴幰偼捝傒傪妎偊傞丅乮挘椡偑庛偡偓傞偲岠偐側偄乯
仠摍広惈廂弅傗摍挘惈廂弅偱偼庴幰帺怣偑廂弅偺掱搙傪捝傒傪姶偠側偄條偵廋惓偱偒傟偽斾妑揑埨慡丅
仠摍挘惈廂弅偱偼嬝挿偑抁偔側傝側偑傜廂弅偡傞偺偱丄摿偵晹暘揑偵彎傫偱偄傞嬝慇堐偑偁偭偨偲偟偰傕曐岇偝傟傗偡偄丅
婋尟惈偵偮偄偰丗
仠摍広惈廂弅偱偼嫮偄廂弅偑摼傜傟傞偺偱崅偄寣埑忋徃偑婲偙傝摼傞丅
庴幰偺妝偝壛尭丗
仠懠摦揑怢挿偱偼惍幰偵懱傪梐偗偰偄傞偩偗偱椙偄偺偱庴幰偼妝偱偁傞丅
仠摍広惈廂弅傗摍挘惈廂弅偱偼庴幰偼擻摦揑偵嶲壛偟側偗傟偽側傜側偄偺偱斾妑揑尦婥偑側偄偲晄岦偒丅
偲偄偆傢偗偱丄庢傝姼偊偢嬝擏憖廲朄偵偮偄偰偺摉弶媈栤偼夝寛偟丄堦曕摜傒崬傫偱棟夝偱偒偨傛偆側婥偑偡傞偺偱偡偑丄價乕僶乕偝傫偼擛壗偍峫偊偱偟傚偆偐丅
Goopon
偆乕傫丄愭偵傕尵偭偨傛偆偵栤戣傪偄偔偮偐姶偠偰偄傞丅傑偢傂偲偮偼嬝擏偵懌偟嶼傗堷偒嶼偼娭學偡傞偺偐側乮屻夞偟乮2乯乯丅傆偨偮傔偼乽嬝朼悗傪姶偠偝偣側偄傛偆偵憖嶌偡傞乿偲偄偭偰傞偗傟偳丄偍偦傜偔偦傟偼柍棟偩傛丅偙傟偼柍帇偟偰偟傑偭偨兞慄堐偑偐傜傫偱偔傞丅屻夞偟乮1乯偺栤戣偩傛丅
傑偢丄傂偲偮傔偺丄嬝擏偺懌偟嶼傗堷偒嶼偺栤戣傪曅偯偗偪傖偍偆丅侾倐偺梷惂傕偐傜傓丄偙偺栤戣偼兛嵶朎偺慜偲屻偱峫偊偰傒側偗傟偽側傜側偄偲巚偆側丅兛嵶朎偺敪壩偲偦傟埲慜偺栤戣偩丅傕偆堦搙丄惍棟偡傞偲丄嬝擏傪嬞挘偝偣傞偵偼兛慄堐偵怣崋乮揹棳乯偑棳傟側偗傟偽側傜側偄丅偦偺堊偵偼兛嵶朎偑敪壩偟側偗傟偽側傜側偄偑丄偦偺兛偺慜偲屻偱偼丄彮乆帠忣偑堘偭偰偔傞丅
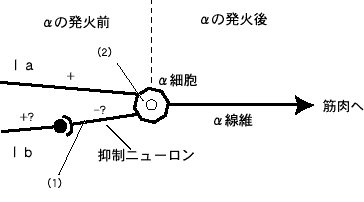
戞俇恾
愭偵兛偺敪壩屻偐傜峫偊傞傛丅偦偺栤戣偲偼丄嬝擏偺婎杮尨懃偼丄怣崋偺桳傞偐柍偄偐丄侾丄侽偟偐側偄丅侾側傜嬝擏偼弅傓偟丄侽側傜弅傑側偄丅
亀捠忢偺悘堄塣摦偵傒傜傟傞崪奿嬝偼丄庡偵偙偺屻幰偺宍幃偵傛傞惂屼傪庴偗偰偄傞丅崪奿嬝偺嬝慄堐偼丄偳傫側巋寖偵懳偟偰傕丄偦傟偑斀墳傪婲偙偡偺偵廫暘側嫮偝偺傕偺偱偁傞尷傝丄巋寖偺嫮偝偵娭傢傝側偔嵟戝偺斀墳傪帵偡丅偡側傢偪崪奿嬝慄堐偼慡偐柍偄偐偺朄懃偵廬偆斀墳傪帵偡丅乮僽儕僞僯僇昐壢帠揟乯亁
亀恄宱嵶朎傪巟攝偡傞乽慡偐柍偐偺朄懃乿
丂恄宱嵶朎偺嫽暠偼丄旕慄宍悢妛偺悽奅偱偄偆僇僆僗揑曄嬋揰傪傕偮丅嫽暠偡傞偐偟側偄偐偱偁傞丅惗棟妛偱偼偙傟傪乽慡偐柍偐偺朄懃乿偲偄偆丅嫽暠惈偺忣曬偑廤愊偟丄恄宱嵶朎偺枌偺揹婥揑暯峵偑偁傞堦揰傪墇偡偲丄偦偺恄宱嵶朎偼堦嫇偵嫽暠傊偲憱傞丅偙偺曄嬋揰傪嫽暠偺鑷抣偲偄偆丅
丂悢昐偺侾倎慇堐偺嫽暠惈揱摫偺憤榓偑偙偺鑷抣傪挻偡偲丄偦偺恄宱嵶朎偼嫽暠偟丄偦偺巟攝嬝偵嬝廂弅傪婲偙偡傛偆側巜椷傪敪偡傞丅偙偺嬝廂弅偵傛傞懌偺挼偹忋偑傝偑丄銯斀幩偲偟偰懆偊傜傟傞丅
丂堦屄偺嬝擏偼丄悢廫偐傜悢昐偺塣摦恄宱嵶朎乮兛塣摦僯儏乕儘儞乯偱巟攝偝傟偰偄傞丅寬忢幰偱偼丄銯斀幩偱嫽暠偟敪壩偡傞塣摦恄宱嵶朎偼丄偦偺嬝偺慡塣摦恄宱偺偍偍傛偦侾侽乣俀侽僷乕僙儞僩偱偁傞丅乮懱偺斀幩偺傆偟偓妛乯亁
偲偄偆偙偲偩偐傜丄偳傫側偵侾倎傗侾倐偺斀墳偑嫮偐傠偆偲丄兛嵶朎偑堦搙敪壩偟偰偟傑偊偽丄偦偺嫮偝偵偼娭學側偔丄堦棩乽侾乿偱乽怣崋偑偁傞乿偲偄偆偙偲偱廂弅偡傞偺偩偐傜廂弅偝偣傞嫮偝偺嵎偵偼側傜側偄傛丅偮傑傝敪壩偟偨屻偱偼懌偟抜傗堷偒嶼偼惉傝棫偨側偄丅
偩偐傜丄goopon偝傫偺偄偆堷偒嶼偑惉棫偡傞偺偼丄兛偺敪壩慜偺栤戣偵側傞偲巚偆丅偟偐偟丄偙偙偵傕栤戣偑偁偭偰丄梷惂僯儏乕儘儞乮1乯偺栤戣偲梷惂僯儏乕儘儞偲兛嵶朎偺娫乮2乯偺栤戣偺擇偮偑偁傞偲巚偆丅
乮1乯偺栤戣
偦傕偦傕梷惂僯儏乕儘儞偲側側傫側偺偩丅
梷惂惈乮壔妛乯僔僫僾僗
梷惂惈壔妛僔僫僾僗偼丄嫽暠惈壔妛僔僫僾僗偲摨條偺曄壔傪婲偙偡偑丄壔妛揱払暔幙偵傛偭偰丄屻晹枌偵撪岦偒偺揹棳偑棳傟偰夁暘嬌傪婲偙偝偣傞偨傔偵丄嫽暠惈偑掅壓偡傞丅偙偺傛偆側壔妛揱払暔幙傪梷惂惈揱払暔幙偲偄偄丄偙偺帪丄屻晹枌偵傒傜傟傞夁暘嬌偺揹埵曄壔傪梷惂惈僔僫僾僗屻揹埵乮俬俹俽俹乯偲偄偆丅偙偺曄壔偼丄梷惂揱払暔幙偵傛偭偰屻晹枌俲僾儔僗偲俠倢儅僀僫僗偺摟夁惈偑崅傑傝丄俲僾儔僗偺暯峵揹埵偐傜俠倢儅僀僫僗暯峵偺曽傊孹偒丄夁暘嬌傪婲偙偟偰偔傞傕偺偲峫偊傜傟偰偄傞丅乮恾夝惗棟妛乯
扙暘嬌乮夁暘嬌乯
惷巭帪偺嵶朎偼丄嵶朎枌偺撪奜偵嵶朎撪傪晧偲偡傞揹埵嵎傪桳偟偰偄傞丅偡側傢偪嵶朎枌偼暘嬌偟偰偄傞偲偄偊傞丅庬乆偺尨場偱偙偺嵶朎枌傪捠偟偰丄奜岦偒偵揹棳偑棳傟傞偲丄暘嬌偑彫偝偔側傞丅偙傟傪扙暘嬌偲偄偄丄嫽暠惈枌偱偼扙暘嬌偵傛傝偱枌揹埵偑尭彮偟丄鑷抣乮椪奅枌揹埵乯偵払偡傞偲妶摦揹埵傪惗偠傞丅乮堛妛戝帠揟乯
傛偔傢偐傜傫偑丄僯儏乕儘儞乮恄宱嵶朎乯偼偦偺恄宱慄堐偱丄懠偺恄宱嵶朎偲愙懕偟偰偄傞丅偦偺愙懕晹埵偼僔僫僾僗乮恄宱愙崌乯偲偄偭偰偄傞丅偦偟偰丄僯儏乕儘儞乮恄宱嵶朎乯偺恄宱慄堐偺愭抂晹偼彫摢偲偄偆嬍偙傠偵側偭偰偄傞丅偙傟傪僔僫僾僗彫摢偲偐僔僫僾僗儃僞儞偲偄偭偰偄傞丅偦偺儃僞儞偺晹暘偼亄偐亅偺揹壔傪偍傃偰偄傞丅亅傪偍傃偨僔僫僾僗彫摢傪帩偮僯儏乕儘儞偑梷惂僯儏乕儘儞丅偁偁丄傗傗偭偙偟乕丅
偦傟偱梷惂僯儏乕儘儞偵偮側偑傞偲丄侾倐偺怣崋偼偳偺傛偆偵曄壔偡傞偐偲偄偆偲丄偙傟偼擇偮峫偊傜傟傞丅堦偮偼壖偵100偺抣偺傕偺偑丄90偖傜偄梷惂偝傟偰10偺抣偵側偭偰偟傑偆応崌丅傕偆堦偮偼姰慡偵斀揮偟偰丄偮傑傝亄偑亅偵揮偠偰亄100偑亅100偲側偭偰偟傑偆応崌丅巹偺帩偭偰偄傞杮偱偼丄偙偺偁偨傝偑偳偆側偭偰偄傞偺偐彂偄偰偄側偄偺偱丄悇應偡傞偟偐側偄丅
傑偢梷惂僯儏乕儘儞偑慜幰偺惈幙傪帩偭偰偄傟偽丄goopon偝傫偺壖愢偼偙偙偱惉傝棫偨側偔側傞丅側偤側傜侾倐偺怣崋偑斀揮偟側偄偺偱丄忢偵亄偺忬懺偱偄傞偐傜偩丅侾倎傪梷惂偡傞偨傔偵偼梷惂僯儏乕儘儞偼亅偵側偭偰偄側偗傟偽偄偗側偄偑丄100偑90梷惂偝傟偰10巆偭偰偄傞丄偦偺10偼丄彫偝偔偼偁傞偑亄偱偁傞丅偟偨偑偭偰侾倎偼亄丄侾倐傕彫偝偄偗傟偳亄偵側傝丄侾倐偺摥偔寢壥偼偡傋偰嬞挘偲側傞丅偙傟傪憖嶌1偺昞偺悢帤偱偄偆偲丄嬞挘偑亄侾側傜抩娚偼亅侾偱偼側偔丄亄0.1偱傢偢偐側嬞挘偲側傞丅偙偆側傟偽goopon偝傫偑昞偺拞偱偄偆梷惂偼婲偙傜側偄丅偡傞偲奺昞偺偡傋偰偵抩娚偑柍偔側傝丄偡傋偰偑嬞挘偟偰偟傑偆偺偩偐傜goopon偝傫偺壖愢偼惉傝棫偨側偔側傞丅
壖愢偑惉傝棫偮偵偼丄偙偺梷惂僯儏乕儘儞偑屻幰丄偡側傢偪姰慡偵怣崋傪斀揮偝偣偨応崌偩偗偱偁傞丅偡側傢偪亄侾倐傪亅侾倐偲偡傞応崌偱偁傞丅偙偺慄偱榑傪恑傔偰峴偔丅
乮2乯
偙偙偱斀揮偑婲偙傞偲偟傛偆丅偡傞偲崱搙偼梷惂僔僫僾僗偲兛嵶朎偺娫偱偺栤戣偵側傞丅戞俇恾偺乮2乯晹暘偩丅偙偙偱偼侾倎偲侾倐偱偺揹棳偺懌偟嶼傗堷偒嶼偑惉棫偟偦偆側婥偑偡傞偑丄偦偆偡傞偲峫偊傜傟傞偺偼兛嵶朎傪敪壩偝偣側偄傛偆偵乽侾倐偑姰慡偵侾倎傪梷惂偡傞乿偲偄偆偙偲偑栤戣偵側傞丅偩偑丄偙偙偱傕goopon愢偼惉傝棫偨側偔側傞丅
侾倎偑丄壖偵亅1偱梷惂偝傟傞側傜偽丄嬝擏傪嫮偔堷偄偰侾倐偵棳傟傞揹棳傪崅傔偰-2丄-3丒丒丒偵偟偰傕丄偡偱偵-侾偱兛嵶朎偼梷惂偝傟偰偄傞偺偩偐傜丄梷惂傪嫮傔偨偲偙傠偱壗傪偟傛偆偲兛嵶朎偼敪壩偟側偄丅嬝擏偼乽慡偐柍偐偺朄懃乿偵偟偨偑偆偐傜偩傛丅兛嵶朎偺鑷抣偵亅侾偱鑷抣偵払偟側偐偭偨偺偩偐傜-2丄-3丒丒丒偲側傟偽丄偝傜偵払偟側偄丅廂弅偝偣傞偨傔偺怣崋偼傛偗偄棃側偄偙偲偵側傞偩偗偩丅偟偨偑偭偰丄憖嶌俀乣俇偺憖嶌偼偡傋偰惉傝棫偨側偔側傞丅
偲偄偆傢偗偱丄goopon偝傫偺壖愢偼偡傋偰偺柺偱斲掕偝傟偰偟傑偭偨丅偱傕丄偙偺寢榑偼撉傫偱偄傞恖偵偼娙扨偵尒偊傞偐傕偟傟側偄偑丄偙偙偵払偡傞偵偼梕堈偱偼側偐偭偨傛丅幚偼嶐擔丄偙傟傪彂偄偨帪揰偱偼丄斲掕偱偼側偔goopon偝傫偺壖愢傪峬掕偟偰彂偄偰偄偨丅偦偺慜偼斲掕丄偦偺慜偼峬掕丄俀揮偳偙傠偐係揮丄俆揮偟偰偄傞偺偩丅堷偭偐偐偭偰偄偨偺偼丄乽侾倐偼丒丒丒偦偺嬝擏偺兛偵梷惂柦椷傪弌偡偺偱丄偦偺嬝擏傪抩娚偝偣傛偆偲偟傑偡丅乿偲偄偆尵梩偩偭偨丅
偳偺夝朥傗惗棟妛偺杮傪撉傫偱傕摨偠傛偆偵彂偄偰偁傞丅傗偼傝惓偟偄偺偐側偲巚偭偰偟傑偭偨丅偦偙偱愭偺傛偆偵暘夝偟偨傢偗偩偑丄偦偙偱嬝擏偲偄偆偺偼傾僫儘僌怣崋偱曄壔偡傞偺偱偼側偔丄僨僕僞儖偱曄壔偡傞偙偲偵婥偯偄偨偺偩丅婥偯偗偽娙扨偩偭偨丅尵梩偑栤戣側偺偩丅-2丄-3丒丒丒偲挘椡偲憹壛偝偣偰備偔偲丄偮偄丄偦傟偵偮傟偰梷惂椡傕憹壛偡傞傛偆偵巚偭偰偟傑偆丅亀兛偵梷惂柦椷亁偼傾僫儘僌偱偩偝傟偰偄傞傢偗偱偼側偔僨僕僞儖側偺偱偁傞丅偩偐傜丄亀兛偵偼侾倎偐傜偺廂弅(+)柦椷偲侾倐偐傜偺抩娚(-)柦椷偑摨帪偵揱傢傝丄偦偺弖娫弖娫偵柦椷偺嵎偟堷偒傪偟偰丄偦偺寢壥廂弅柦椷偑桪惃偱偁傟偽廂弅偟丄抩娚柦椷偑桪惃偱偁傟偽抩娚偟傑偡丅亁偺寁嶼傕僨僕僞儖偺寁嶼偲偲傜偊傞傋偒偩偭偨偺偩丅
丂
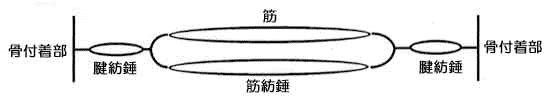
嬝朼悗偺旈枾
偡偱偵丄goopon偝傫偺壖愢偑惉傝棫偨側偄偙偲傪棫徹偟偨偺偩偐傜丄偙偙偱捛媮傪偆偪愗傞傋偒偐傕偟傟側偄丅傑偨丄懡偔偺恖払偵偼巹払偑婘忋偺嬻榑傪傕偰偁偦傫偱偄傞傛偆偵尒偊傞偐傕偟傟側偄丅偟偐偟丄巹偼傑偩偙偺捛媮傪懕偗傞偮傕傝偱偄傞偺偩丅偦傟偼丄goopon偝傫偺壖愢偺姰晢柍偒傑偱偵偆偪攋傞偨傔偱偼側偄丅偙偙偐傜怴偟偄壴傪嶇偐偣傞偮傕傝偩偐傜丅
榖傪彮偟旘偽偡偑丄婽堜愭惗偑愢偄偨島媊偺拞偵偼丄島媊傪傑偲傔傞嵺偺曇廤偺偢偝傫偝偐傜丄懡偔偺撲偺晹暘偑偁傞丅偙傟偐傜弎傋傞乽嬝朼悗乿偵偮偄偰偺島媊偼丄偍偦傜偔撲偺拞偱傕嵟戝偺傕偺偩丅
亀嬝擏偼捠忢丄婲巒晹偲掆巭晹偲偑偁偭偰丄偦偺拞娫偵嬝暊偑偁傞偺偱偁傞偑丄偙傟偵偼墶偺慄堐偲廲偺慄堐偲偑偁偭偰丄嬝擏偑廂弅偡傞偲偄偆帠偼丄偙傟傜偑朿挴偡傞偺偱偁傞丅偮傑傝墶幦偺慄堐偑墶偵僗儕僢僾偟偰偼傒弌偟偰丄偁偨偐傕廂弅偟偨偐偺傛偆偵尒偊偰姶偠傜傟傞偺偱偁傞丅偙傟偑懎偵偄傢傟偰偄傞廂弅偟偨偲偄偆忬懺側偺偱偁傞丅
廲偺慄堐偱偁傞嬝朼悗偼壗偺偨傔偵偁傞偺偐偲偄偆偲丄堦庬偺斀幩慄堐偺栶傪柋傔丄暔傪揱払偡傞偲偄偆嶌梡偑偁傞丅偮傑傝嬝朼悗偑巋寖傪庴偗傞偲朿挴偡傞丄偲偄偭偰傕丄偙偺傑傑朿挴偡傞帠偼弌棃側偄偐傜弅傑傞丅朿挴偡傞帠偲弅傑傞帠偺擇偮偱嬝擏偼廂弅偡傞偺偱偁傞丅僗儕僢僾偡傞帠偲朿挴偡傞偲偄偆帠偼摨偠傛偆側堄媊偵巚傢傟傞偑偙傟偼堘偆丅嬝朼悗偲偄偆偺偼怢偽偟偨偲偒偵偺傒擼偲偐塣摦偺媮怱巋寖偵側偭偰揱傢偭偰偄偔偺偱偁傞丅偩偐傜嬝朼悗偑怢傃傞偲偄偆帠偼丄偦偺嫽暠偑拞悤偵岦偭偰揱払偝傟側偗傟偽側傜側偄嶌梡傪昁梫偲偟偨偲偒偵偺傒朿挘偡傞偺偱偁傞丅墶幦偺慄堐偑墶偵僗儕僢僾偡傞偺偲偼偦偺惈幙偼堘偆偺偱偁傞丅
嬝朼悗偺朿挴偼擼傪旕忢偵妎惲偡傞嶌梡偑偁傞丅嬝朼悗偑惓忢偵側偭偰僒乕僢偲怢傃偒偭偨偲偒偵偼斀墳傪慡偔婲偙偝側偄丅偙傟摍偼嬝擏偺惗棟偱偁傞偐傜傛偔棟夝偟偰偍偐側偄偲挷惍忋偵墬偰崲傞丅偱偼昦婥傪恌傞偲偒偵偳偆恌傞偺偐偲偄偆偲幦柾條偵側偭偰偄傞僗儕僢僾偟偰偄傞慄堐摍傪挷傋偰傒偰傕堄枴偑側偄偺偱偁傞丅
娭愡偑堎忢傪婲偙偟偨傝丄摦偐側偔側偭偨傝偟偨偲偒偵偼墶偵僗儕僢僾偡傞慄堐偱偼側偄偺偱偁傞丅廲偺嬝朼悗側偺偩丅偙偺尨棟偑暘偐傜側偄偺偱偼偄偔傜挷惍傪壛偊偰傕帯傜側偄丅偙偺尨棟偑傛偔棟夝偝傟偰偄傞偲娭愡摍偺摦偐側偔側偭偨傕偺偺挷惍偼旕忢偵傗傝堈偄丅
嬝朼悗傪挷傋傞偲偒偼墶偵抏偄偰挷傋傞丅巜偺旂晢偺姶妎偑塻晀側恖偼寉偔墶偵抏偄偰偄傞偲偦偺嬞挘偵怗傟傞偺偑傛偔傢偐傞丅偙偺嬞挘偼慡慄堐偑偡傞偺偱偼側偔偰堦晹偺嬝朼悗慄堐偑嬞挘偡傞偺偱偁傞丅偮傑傝墶偺幦柾條偺慄堐偼戝側傝彫側傝慡懱偑墶偵僗儕僢僾偟偰偍傞偑丄嬝朼悗偺応崌偼椡偺擖梡検偲偐偁傞偄偼巋寖偺揱払偺堦偮偺曽朄偲偟偰庬乆嶨懡偵嬞挘偡傞杮悢偑堘偆偺偱偁傞丅偦傟偵傛偭偰偄傠偄傠偺帠偑惗偠傞偲偄偆惈幙偑偁傞偺偱偁傞丅
嬝朼悗偼怢傃偒偭偰偄傞偲偒偼壗偺嶌梡傕偟偰偄側偄丅摦偐偟偨偲偒傕摨條偱偁傞丅弖娫偵僷僢偲怢偽偡丅偙偺僷僢偲怢偽偡弖娫偺巋寖偩偗偵墳偠傞嶌梡偑偁傞偺偱偁傞丅偁傞偄偼枖僌僂乕僢偲堷偭傁偭偰怢偽偟偰偍偄偰僷僢偲僔儑僢僋傪壛偊傞偲墳偠傞嶌梡偑偁傞丅懄偪怢斀幩偱偁傞丅嬝朼悗偲偄偆偺偼孅嬝懁傛傝怢嬝懁偺曽偵懡偔敪払偟偰偄傞丅丒丒丒乮婡娭帍26崋乽巋寖偺婎慴尨棟乿乯亁丂
撉傫偱暘偐傝傑偡偐丅嬒惍朄偺擄夝側暥復拞丄偙傟傎偳傓偪傖偔偪傖偱棟夝偵嬯偟傓暥復偼側偄偺偱偡丅偙傟偼婽堜愭惗偺榖偺巇曽偑偁偭偪旘傃偙偭偪旘傃偟偰丄傑偨嵟弶偵栠傞丄偲偄偆曇廤幰媰偐偣偺傕偺偱偁傞忋偵丄傢傟傢傟偺傛偆側曇廤幰傕慺恖丄暥復姶妎偁傑傝側偄偆偊丄堛妛抦幆偵傕惛捠偟偰偄傞傢偗偱偼側偄丅巹傕娷傔偰丄嬒惍巘偲偄偆傕偺偼帺枬偠傖側偄偑丄摢傪巊偭偰怘偭偰偄傞偺偱偼側偄丅庤傪巊偭偰偄傞偺偩偐傜丄棟榑揑抦幆偵偮偄偰偼庛偄丅偮傑傝堛妛偺抦幆偺偁傑傝側偄恖偨偪偑丄乽嬝朼悗乿偲偄偆偙傟傑偨擄夝偺栤戣傪傑偲傔傛偆偲偡傞偺偩偐傜丄傑偲傔傞嵺偵億僀儞僩傪偼偢偟偰偟傑偭偨偺偱偼側偄偐偲巚偆丅偱傕丄偛怱攝側偔丄偙傟偐傜巹偑愢柧偡傞偲偙傠傪撉傔偽丄偙偺抐曅揑側婰弎偺堄枴偑偐側傝暘偐偭偰偔傞偼偢偱偡丅偦偺偒偭偐偗偲側偭偨偺偑goopon偝傫偺壖愢側偺偱偡丅
goopon偝傫偼丄側偤嬞挘偟孹幬傪婲偙偡嬝擏偑娚傓偺偐偵偮偄偰丄偍偦傜偔嬒惍巘偱偼扤傕傗偭偨偙偲偺側偄曽朄偱丄偦偺撲偵挧傫偩偺偩丅偟偐偟丄偦偺棟榑棫偰偵偼岆傝偑偁傝偦偆偵傒偊傞丅側偤岆偭偨偺偐丄偦偺尨場傪暿偺妏搙偱傒偰傒傞偲丄埲奜傗埲奜丄崱傑偱嵟戝偺撲偲偝傟傞偺偑丄夝偗偰偟傑偭偨丅偙傟偵偼丄偲傝偔傫偩巹偑傃偭偔傝偟偨丅
岆傝偦偺侾丗 儈僢僔儞僌儕儞僋
崱堦搙丄巹偺斀榑偑娫堘偭偰偄偰丄goopon偝傫偺壖愢偑惓偟偐偭偨偲偡傞丅偦傟偱傕丄偙偺愢偵偼擄揰偑偁傞丅偦傟偼丄偡傋偰偑goopon偝傫偺尵偆偲偍傝偩偭偨偲偟偰丄偮傑傝丄侾倎傪梷惂偡傞偨傔偵丄乽乮憖嶌俇乯偑抩娚嶌梡偑堦斣嫮偄乿偲偄偆偺偑丄壖偵惓偟偔偰傕丄憖嶌傪傗傔偰偟傑偊偽尦偺傕偔偁傒偵側傞丅
偳偆偄偆偙偲偐偲偄偆偲丄壖偵嬝擏偵嬅傝偑偁偭偰丄偄偔傜偐偺嬞挘偑偁傞偲偡傞丅傑偁丄偦傟傪10偲偟傛偆偐丄偮傑傝侾倎偐傜偼10偲偄偆嬞挘偺儗儀儖偑兛偵敪偣傜傟偰偄傞丅偦偺忬懺偱丄亅80偲偐亅90偲偐偵堷偭挘傞傢偗偩丅偱偼丄偦偺堷偭挘傞椡傪尦偺侽偵傕偳偟偨傜丄侾倎偲侾倐偺捈愙揑側娭學偑柍偄偺偱乮擇師揑偵偼偁傝偦偆偩偑乯丄堷偒嶼偑惉棫偟偦偆偵側偄丅偩偐傜丄侽偵栠偟偨傜丄尦偁偭偨10偺嬞挘偑傑偨偁傜傢傟偰丄嬝擏偼嬞挘偟偨傑傑偲側傞丅侾倎偲侾倐偺娭學偐傜偱偼丄侾倎偺嬞挘傪夝偔偨傔偺儊僇僯僘儉偑懚嵼偟側偄偺偩丅偩偐傜丄嬝擏偑娚傓偙偲偵偼側傜側偄丅
偲偼尵偊丄嬝擏憖廲偱巊偆僥僋僯僢僋偺堦偮偺嬝擏傪堷偒怢偽偡曽朄偱尰幚偵娚傫偱偄傞偙偲傕妋偐偩丅偱偼丄壗偑嬝擏傪娚傑偣偰偄傞偺偐丅妋偐偵嬝擏傪堷偗偽丄偦偺帪揰偱嬞挘偼柍偔側傞丅偱傕丄傗傔傟偽傑偨尦偵栠傞丅娚傓偨傔偺儊僇僯僘儉偲偄偆偐丄偦偺娭學偲偄偆傕偺偺愢柧偑側偄丅侾倎偺敪壩偵塭嬁傪梌偊傞壗偐偑偁傞偼偢偩丅偦傟偼丄侾倎偲侾倐偺娭學偩偗偱偼嬝擏偺抩娚偺尨棟傪棟榑壔偟傛偆偲偟偰傕丄偙偺擇偮偺娭學偩偗偱偼柍棟偑偁傞偺偩丅侾倎偲侾倐傪偮側偖壗偐偑昁梫偵側傞丅 偪傚偆偳椶恖墡偲恖娫偲偺拞娫偵偁偭偨偲壖憐偝傟傞摦暔偺幐傢傟偨娐嵔乮儈僢僔儞僌儕儞僋乯偺傛偆側傕偺偑丅
goopon偝傫偼側偤偙偺擇偮偺娭學偵峣偭偨偐偲偄偆偲丄懠偺恄宱偵庤傪峀偘傟偽丄娭學偼婔壗媺悢揑乮屆偄偹乯丄梫偡傞偵偳傫偳傫娭學偑暋嶨壔偟丄忣曬張棟偑暋嶨壔偟偡偓偰丄偲偰傕庤偵晧偊側偔側傞偐傜丄偙偺擇偮偺娭學偵峣偭偨偺偩偲巹偼巚偆傛丅偩偐傜暋嶨側栤戣偺棈傓兞傪傢偞偲偼偢偟偨偺偩丅栤戣偼偙偙偵偁傞丅偦偟偰偙傟偼屻夞偟乮1乯偺栤戣偱傕偁傞丅
岆傝偦偺俀丗兞傪柍帇偡傞側
嬝擏傪廂弅偝偣傞兛偵峴偔怣崋偼侾倎傗侾倐埲奜偵傕偁傞丅壓偺恾傪尒偰梸偟偄丅戝曄側悢偺恄宱偑嬝擏偵偼擖偭偨傝弌偨傝偟偰偄傞丅
戞俈恾
2.嬝朼忇偺恄宱巟攝乮恾夝惗棟妛傛傝乯
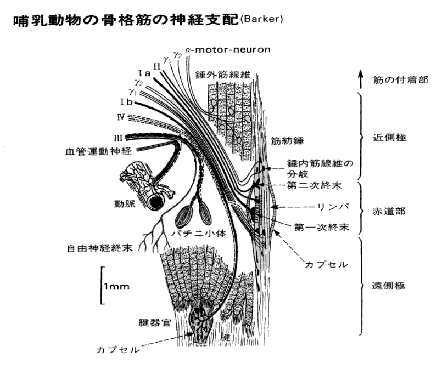
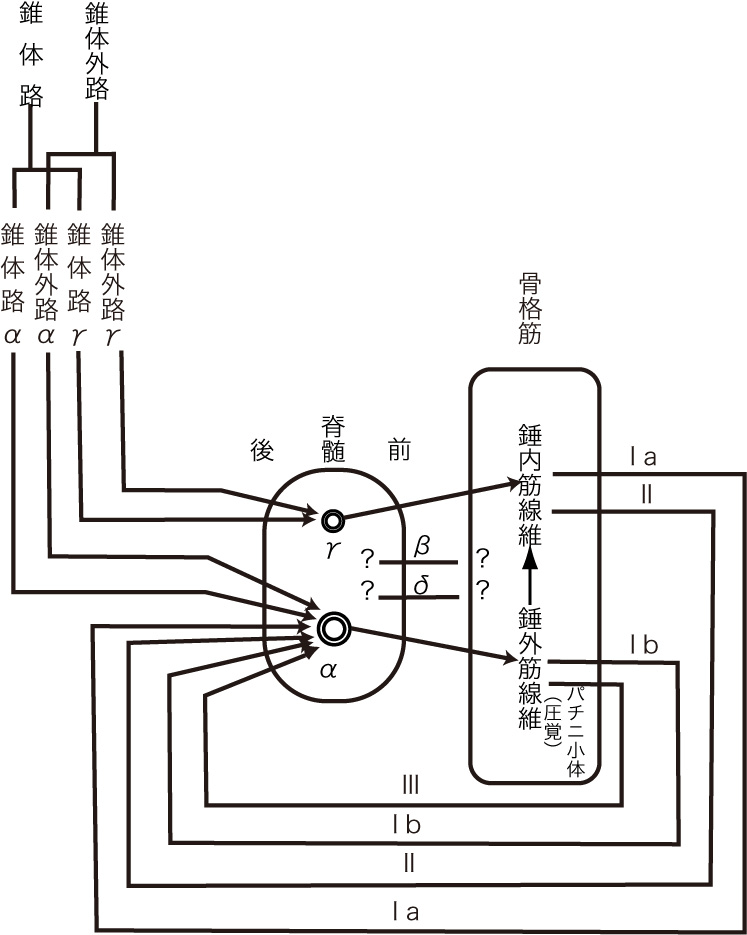
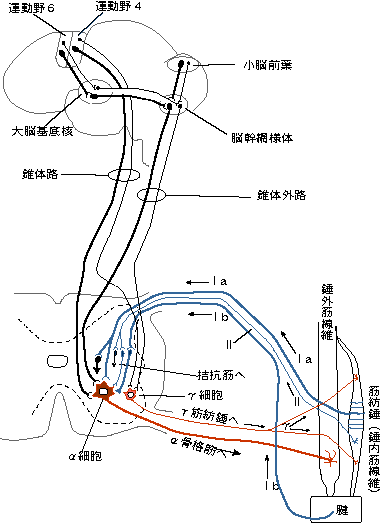
崪奿嬝偐傜愐悜傊忋峴偡傞姶妎恄宱慄堐偵偼丄侾倎(group 侾a fiber),侾倐(group 侾b fiber)丄俀(group俀fiber)丄俁(group俁fiber乯偍傛傃係孮慄堐(group係fiber)偺5孮偑偁傞丅la偍傛傃lb孮慄堐偼A兛慄堐偵丄俀孮慄堐偼A兝偍傛傃A兞慄堐偵丄俁孮慄偼A兟慄堐偵丄係孮慄堐偼C慄堐偵憡摉偡傞丅偙偺暘椶偼嬝姶妎恄宱偵尷偮偰梡偄傜傟偰偄傞丅側偍丄崪奿嬝偺怢揥傗廂弅偺忬懺丄斾妎丄怳摦偺姶妎丄捝妎側偳傪拞悤偵揱偊傞偺偼偙傟傜偺恄宱偵傛偭偰峴傢傟偰偄傞丅
忋婰偺暥彂偼嬝擏偐傜弌偰偄偔恄宱丄偮傑傝傾僂僩僾僢僩偡傞恄宱偵偮偄偰弎傋偨傕偺偱偁傞丅僀儞僾僢僩偼弎傋傜傟偰偄側偄丅偦偙偱擖椡偲弌椡傪恾偵偡傞偲師偺傛偆偵側傞丅
 丂
丂
戞俉恾
偡偛偄偹丄偨偭偨堦杮偺嬝慄堐偵偙傟偩偗偺忣曬偑弌擖傝偟偰偄傞丅偟偐傕丄傑偩係孮偺慇堐偼徣棯偟偰偄傞偺偩丅偙偺拞偱傕偭偲傕拲栚偵抣偡傞偺偑兞偺摥偒側偺偩傛丅偦偙偱兞慄堐乮悗撪嬝慄堐丒嬝朼悗乯偲偼壗偐偲偄偆偙偲偵側傞丅
偙傟偑丄傢偐傝偵偔偄奣擮側偺偩偗傟偳丄偡偽傜偟偄愢柧偑僀儞僞乕僱僢僩偺拞偵偁偭偨偺偱堷梡偝偣偰傕傜偭偨丅埲壓偺愢柧偼懌愳偝傫偲偄偆棟妛椕朄巘偺曽偺儂乕儉儁乕僕傛傝揮嵹丅嬝朼悗偺婡擻夝朥偺峫嶡側偺偩偗傟偳丄幚偵暘偐傝傗偡偄丅尃埿傪庢偭暐偆偲暘偐傝傗偡偔側傞偲偄偆岲椺偺傛偆側婥偑偡傞丅巹偑夝愢偡傞傛傝傢偐傝傗偡偄偺偱巊傢偣偰傕傜偭偨丅奆偝傫傕惀旕堦搙偼朘傟傞傋偒偱偡乮http://homepage.mac.com/kazz2000/ref/reflex.html乯丅儂乕儉偼http://homepage.mac.com/kazz2000/丅偟偐偟丄巹傕抪偢偐偟偄丅嬝擏傪枅擔挷惍偟偰偄傞偺偵丄偙偆偄偆尒棊偲偟傪偟偰偄偨偲偼丅悽偺拞峀偄丄偦偟偰僀儞僞乕僱僢僩偰偲傫偱傕側偄偲偙傠偩偲偮偔偯偔巚偆側丅偁丄偙傟偼梋択偩偹丅
兞偲偄偆偺偼悗撪嬝慄堐傪廂弅偝偣傞摥偒偑偁傞丅偙傟偑億僀儞僩丅丂
丂丒丒丒嬝朼悗偼丄偨偔偝傫偁傞嬝慄堐偺拞偵晹暘揑偵懚嵼偡傞偺偱丄嬝偑怢偽偝傟偨偲偒偵偼巇曽側偔堦弿偵嬝朼悗傕怢偽偝傟傑偡偑丄兛塣摦僯儏乕儘儞偵傛偭偰嬝偑弅傓偲偒偼丄忋恾偺傛偆偵嬝朼悗偼偨傞傫偱偟傑偆傢偗偱偡丅偦傟偱偼儎僶僀偱偡丅
丂偦傟偱偼丄偙偺婋尟傪偳偆旔偗傟偽傛偄偐丒丒丒丄偦傟偑偙偺嬝傪抁偄傑傑堦掕偵曐偮乮嬝朼悗偺姶搙挷愡婡峔乯側傢偗偱偡丅夝寛朄偼丄嬝偑抁弅堟偱傕嬝朼悗偑挘椡傪庴偗傞偙偲偑弌棃傟偽傛偄傢偗偱偡丅丒丒丒偲偄偭偰傕偳偆偡傟偽傛偄偐丅僂乣儞丂丂偲擸傫偩偁偘偔嬝朼悗傪廂弅偝偣傞傢偗偱偡偹丅壗屘偐丠丂傑偨傕傗恾傪梡偄偰愢柧偟傑偡丅
偙偙偱怴偟偔丂兞偲尵偆偺偑偱偰偒傑偟偨丅偙傟偼丄兞塣摦僯儏乕儘儞偲偄偭偰丄摥偒偼兛塣摦僯儏乕儘儞偲偲偰傕傛偔帡偰偄傑偡丅兛塣摦僯儏乕儘儞偑嬝乮悗奜嬝乯傪弅傑偣傞偺偵懳偟丄兞塣摦僯儏乕儘儞偼嬝朼悗乮悗撪嬝乯傪弅傑偣傞傢偗偱偡丅壗搙傕偔偳偔尵偄傑偡偑丄兛傗傜兞傗傜侾a,侾b側偳偲偄偆傕偺偼丄婎杮揑偵堦偮偺摥偒偟偐偟傑偣傫丅
丂暥專傪撉傫偱傒傞偲丄乬兞塣摦僯儏乕儘儞偼丄嬝朼悗偺姶搙傪挷愡偡傞偨傔偵摥偔丅乭偲偄偭偨偙偲偑彂偄偰偁傝傑偡偑丄偍偍偘偝丒暣傜傢偟偄偙偲恟偩偟偔丄僕儍儘偵揹榖偟側偗傟偽側傝傑偣傫丅
丂兞塣摦僯儏乕儘儞偲偄偆傕偺偼丄嬝朼悗傪廂弅偝偣傞偙偲偟偐偟傑偣傫丅偦偺寢壥丄嬝朼悗偑姶偠傗偡偔側傞偺偱偡丅乮兞塣摦僯儏乕儘儞偼丄姶搙偑傛偔側傞曽岦偵偟偐摥偒傑偣傫丅姶搙傪棊偲偟偼偟側偄偺偱偡丅偦傟傪挷愡偲尵偆側傫偰丄偪傚偭偲橖枬偱偡偹丅
傇偭偪傖偗偨榖丄惗棟妛傗傜夝朥妛偺杮傪撉傫偱丄崱堦棟夝偱偒側偄偙偲偑丄偙偺恾偱傗偭偲堄枴偑暘偐偭偨偺偩丅偦傟傑偱偼嬝朼悗偼扨側傞挘椡偺僙儞僒乕偺婡擻偲偟偐偲傜偊偰偄側偐偭偨丅壗偱嬝朼悗偑帺傜弅傑側偗傟偽側傜側偄偺偐棟夝偱偒側偐偭偨丅偦傟偼丄忋偺恾偱乽姶搙偠傗偡偔乿偟偰偄傞偺偩偲暘偐偭偨丅傑偩丄棟夝偱偒側偄恖偑偄傞偐傕偟傟側偄偐傜丄師偺椺偱愢柧偟傛偆丅
偨偲偊偽恾偺傛偆偵侾俉侽搙偺壜摦堟偑偁傞娭愡偑俋侽搙偵嬋偑傞偙偲傪峫偊偰傒傞丅
 戞俋恾
戞俋恾戞堦抜奒
兞嵶朎偲兛嵶朎偵忋埵偺丄偙偺応崌偼擼偩偗傟偳傕丄偦偙偐傜俋侽搙偵嬋偘傠偲尵偆巜椷偑偔傞丅偙偺巜椷偼丄栠偭偰戞俉恾傪尒偰傕傜偊偽傢偐傞偑丄悕懱楬偐傜傕悕懱奜楬偐傜傕偔傞丅偝傜偵偛偰偄偹偄偵丄兛慄偵偲兞慄堐偺擇偮偑偁傝丄崌寁係杮偺慄堐傪捠偠偰巜椷偑愐悜偵摓払偡傞傢偗偩丅
愐悜偱偼悕懱楬偐傜偒偨兛慄堐偺巜椷偲悕懱奜楬偐傜偒偨兛慄堐偺巜椷傪兛恄宱嵶朎偱庴偗偰丄悕懱楬偲悕懱奜楬偺兞慄堐偺巜椷偼兞恄宱嵶朎偱庴偗傞丅
俋侽搙偲偄偆埵抲偼丄嬝擏偵偼傢偐傜側偄偐傜丄偲傝偁偊偢嬋偑傞曽岦偵嬞挘偡傞傢偗偩丅
戞擇抜奒
偦偺帪丄嬝朼悗乮悕撪嬝慄堐乯傕銯朼悗乮悕奜嬝慄堐乯傕堦弿偵弅傓傢偗偩偑丄偳偙傑偱弅傓偐偼嬝擏偵偼敾抐偱偒側偄丅偦偙偱嬝擏偼偳傫偳傫弅傫偱備偔丅偡傞偲丄偁傞帪揰傑偱丄偙偺応崌俋侽搙偩偗傟偳丄偦偺帪揰偱嬝朼悗乮悕撪慄堐乯偼丄愭峴偟偰弅傓偺傪傗傔偰偟傑偆丅兞偺塭嬁偩丅
戞嶰抜奒
偦偺寢壥丄嬝朼悗傛傝側偍嬝擏偼弅傕偆偲偡傞偲丄嬝朼悗偼偦傟傪庴偗偰偨傞傒偡偓偰偟傑偆丅偦偙偱敪壩傪傗傔偰偟傑偆丅偡傞偲兛嵶朎傊峴偔侾a偺拞偺揹棳偑侽偵側偭偰丄嬝擏傪弅傔傛偆偲偡傞椡偑懚嵼偟側偔側傞丅偦偙偱嬝擏偼弅傓偺傪傗傔偰丄俋侽搙偺埵抲偵偲偳傑傞偙偲偵側傞丅
偳偆偱偡丅側傫偲岻柇側夞楬丅帺慠偼慺惏傜偟偄丅僽儔儃乕偲巚偭偨丅偟偐偟壗偩傛柧抭孨丅偙傟偵偼棊偲偟寠偑偁偭偨丅
偝偭偒嬝擏偼弅傓偟偐側偄丅桳傞偐柍偄偐偺朄懃偵廬偆偲尵偭偨丅偲偡傞偲丄嬝朼悗傕僨僕僞儖偱弅傓傢偗偩偐傜丄乽偙偺応崌俋侽搙偩偗傟偳丄偦偺帪揰偱嬝朼悗乮悕撪慄堐乯偼丄愭峴偟偰弅傓偺傪傗傔偰偟傑偆丅兞偺塭嬁偩丅乿側傫偰暘偐偭偨傛偆偵彂偄偨偗傟偳丄偙傟偼婲偙傜側偄丅嬝朼悗偑傾僫儘僌偱丄偨偲偊偽俆儃儖僩乮偙傫側偵崅偔側偄偑乯偱俆侽僷乕僙儞僩丄俇儃儖僩偩偲俇侽僷乕僙儞僩弅傓偲偄偆偙偲偼側偄丅僨僕僞儖側偺偩偐傜丄嬝朼悗偩偭偰弅傓偐弅傑側偄偐偩丅
偆乕傫丄傑偨擸傫偱偟傑偭偨丅杮摉偵偙偺栤戣傪峫偊傞偲偒偼丄偙偺楢懕側偺偩丅傗偭偲夝偗偨偲巚偭偨傜丄巚峫偺夁掱偵棊偲偟寠偑偁偭偰丄堦偮屻栠傝偟偰偟傑偆丅
偦偙偱傑偨丄懌愳偝傫偵偛搊応偹偑偄傑偡
偠傟偭偨偄偺偱偼傗偔愢柧偟偰偟傑偄偨偄偺偱偡偑丄偦偺慜偵堦偮幙栤傪偟傑偡丅崱傑偱偺榖傪摜傑偊偰丄塣摦乮嬝廂弅乯傪偡傞偨傔偵偼丄壗偑偳偆側傟偽傛偄偱偟傚偆偐丒丒丒丠丂摎偊偼丄乬兛塣摦僯儏乕儘儞偑敪壩偡傞乮怣崋傪憲傞乯乭偱偡丅偲偄偆傛傝偼傓偟傠丄兛塣摦僯儏乕儘儞偑摥偔偙偲側偟偵偼嬝廂弅偼弌棃側偄偲尵偭偨曽偑傛偄偺偐傕偟傟傑偣傫丅傑丒丄偳偪傜偵偟傠兛塣摦僯儏乕儘儞偑摥偗偽傛偄傢偗偱偡丅椻惷偵峫偊傟偽偐傫偨傫偱偟傚丒丒丒兛塣摦僯儏乕儘儞偼嬝傪廂弅偝偣傞偲偄偆偨偭偨堦偮偺巇帠偟偐偟側偄偺偩偐傜丒丒丒乮僋僪僀丠乯丅
偙偙偱丄婎慴棟夝偺偲偙傠偱弌偰偒偨悕懱楬丒悕懱奜楬偑搊応偟傑偡丅偲偄偭偰傕摝偘崢偵側傜側偄偱壓偝偄丄娙扨偱偡丅悕懱楬丒悕懱奜楬偑丄塣摦傪峴偆偨傔偺擼偐傜偺怣崋偺宱楬偩偲偄偆偙偲偼慜偵傕愢柧偟傑偟偨偑丄偦偺偳偪傜傕丄峴偒拝偔愭偼愐悜慜妏偵偁傞兛嵶朎乮兛塣摦僯儏乕儘儞偺嵶朎乯偱偡丅偮傑傝嬝傪廂弅偝偣傞偺偱偡丅偟偐偟悕懱奜楬偵偍偄偰偼丄偍側偠偔愐悜慜妏偵偁傞兞嵶朎偵傕怣崋傪憲傝傑偡丅乮壓恾偺傛偆偵乯
悕懱奜楬偵偍偄偰丄側偤兞嵶朎偵傕怣崋傪憲傜側偗傟偽側傜側偄偺偐媈栤偵巚偄傑偣傫偐丠丅嬝廂弅傪偡傞側傜兛嵶朎偵偩偗怣崋傪憲傟偽偄偄偺偵丒丒丒偨偟偐丄兞塣摦僯儏乕儘儞偭偰嬝朼悗傪廂弅偝偣傞儌僲偱偡傛偹丅丒丒丒幚偼丄嬝妶摦傪妸傜偐偵偡傞偨傔偵兞嵶朎偵傕怣崋傪憲偭偰偄傞偺偱偡丅偲尵偭偰傕夝傜側偄偲巚偆偺偱丄枅搙偺傛偆偵恾傪梡偄偰愢柧偟偨偄偲巚偄傑偡丅
兞塣摦僯儏乕儘儞偑摥偔偲丄嬝朼悗偑廂弅偟傛偆偲偟傑偡偑丄偦偺慜偵嬝朼悗偵挘椡偑摥偒傑偡丅挿偝偼曄傢傜側偄偺偵帺暘偩偗弅傑傠偆偲偟偨僶僣偱偡偹丅偦偆偡傞偲偳偆側傞偐丠憗偔師儁乕僕傊丒丒丒丅
僇儞偑偄偄恖偼傕偆夝傞偱偟傚偆乮丒丒丒夝傜側偄恖偺僇儞偑偑埆偄偺偐側丒乯丅偱偼丄壓恾丅丒丒丒桭払側偔偟傑偡偹丒丒丒丅

僂乕儞側傫偐慜偵傕傗偭偨偙偲偑偁傞傛偆側丒丒丒丅嬝朼悗偺怢挘巋寖丒侾a丒兛丒嬝廂丒丒丒丅偦偆偱偡両両両乮丠乯怢挘斀幩偱偡丅偟偐偟丄偙偙偱怢挘斀幩偑婲偙傞偲壗偐僩僋傪偡傞偺偩傠偆偐丒丒丒僴僀偟傑偡丅
丂帩懕揑廂弅偼悘堄揑乮偮傑傝堄幆壓偭偰僐僩乯偵峴傢傟偰偄傑偡丅偭偰偙偲偼丄塣摦傪婲偙偦偆偲巚偭偰偄傞娫偢偭偲悕懱楬丒悕懱奜楬偵偼怣崋偑棳傟偰偄傞傢偗偱偡偹丅偦偆偡傞偲丄兛塣摦僯儏乕儘儞偼傕偪傠傫兞塣摦僯儏乕儘儞偵傕忢偵怣崋偑憲傜傟偰偄傞傢偗偱偡丅偡傞偲丒丒丒丅

偙傟傕偳偙偐偱尒偨偙偲偁傝傑偡偹丅2乯嬝傪抁偄傑傑堦掕偵曐偮偺偲偙傠偱偡丅
偝偰丄忋恾偺塃偺忬懺偐傜兞嵶朎偵怣崋偑棃偨傜偳偆側傝傑偡偐丒丒丒丠傕偆傢偐傞偱偟傚偆丒丒丒傢偐傜側偐偭偨傜丄慜儁乕僕偺壓偺恾偐傜尒捈偟偰壓偝偄丅兞偑摥偗偽摥偔傎偳丄嬝偼偳傫偳傫廂弅乮抁弅乯偟偰偄偔傢偗偱偡丅悕懱奜楬偵偍偄偰兞嵶朎偵怣崋傪憲傞偺傕偡偰偨傕偺偠傖側偄偱偟傚偆丅偲偄偆傛傝昁梫偱偡偹丅恖娫偑塣摦傪峴偆偲偒傑偢兞嵶朎傊偺怣崋偑崅傑傝傑偡丅偦偆偡傞偲丄屻偐傜兛塣摦僯儏乕儘儞偑嬝廂弅傪偝偣傞嵺偵偨偡偐傞乮嬝廂弅傪墌妸偵峴偊傞乯偲偄偆傢偗偱偡丅
傕偆堦搙悕懱楬丒悕懱奜楬偺宱楬傪傑偲傔偰偍偒傑偡丅
寢嬊兛偑摥偗偽嬝廂弅偡傞傢偗偱偡丒丒丒傢偐偭偨丠
傑傞偱僐儞僺儏乕僞偩傛丅僐儞僺儏乕僞偑偐偗嶼偡傞偲偒偼丄懌偟嶼傪孞傝曉偟傪崅懍偱傗傞偺偩丅椺偊偽丄俀亊俆偼俀傪俆夞偨偟偰丄儗僕僗僞乕偑俆傪寁應偟偨傜乽傗傔乿偲偄偄丄僒僽儖乕僠儞偐傜儊僀儞儖乕僠儞偵栠傞偙偲傪偡傞丅
戞10恾
偲偄偆偦傟傪摨偠偩丅偙傟傪崅懍偱傗偭偰偄傞偺偩偹丅偨傇傫丄儁儞僥傿傾儉侾侽侽乮係偠傖側偄傛乯偺10,000僊僈僿儖僣偖傜偄偺僗僺乕僪偱丅
側傫偱偩傠偆丅曄偐傕偟傟側偄偑僗僺乕僪偺堊偩偲巚偆丅僔儏儚儖僣僱僢僈乕偺僞乕儈僱僞乕偺榬偺擏傪偼偖偲丄僔儕儞僟偺怢傃弅傒偱榬偑嬋偑偭偨傝怢傃偨傝偟偰偄偨丅偨傇傫桘埑偱摦偔偺偩偲巚偆偑丄偦偺桘傪儌乕僞偱弌偟擖傟偡傞偙偲傪峫偊傞偲丄偁傑傝憗偔偼摦偗側偄丅杮揷偲偐SONY偱儘儃僢僩傪嶌偭偨偗偳丄儌乕僞乕偱偼恖娫偺傛偆偵慺憗偄摦偒偼偱偒側偄丅偙傟偲斾傋傟偽丄偨偩弅傓偩偗偟偐偟側偄嬝擏偺曽偑弖娫偺摦嶌側偳偺摦偒傪嶌傝弌偟傗偡偄偺偱偼側偄偐偲巚偆偹丅
婡摦愴巑僈儞僟儉偺拞偱傾儉儘偼嫄戝儘儃僢僩偺傛偆側儌價儖僗乕僣乮儘儃僢僩宆愴幵乯傪拝偰乮忔偭偰乯妶桇偡傞丅傾儉儘偼媽恖椶偲偼堘偭偨怴恖椶偱偁傞丅塅拡僗僥乕僔儑儞偵惗傑傟堢偭偨娐嫬偑斵傪曄偊偨丅斀幩恄宱偑媽恖傛傝椙偄偺偱偁傞丅偩偐傜斵偑忔傞僈儞僟儉偼嫮偄丄偲偄偆愝掕偩偭偨丅偱傕丄偙傟偼柍棟偩偹丅側偤側傜僒乕儃儌乕僞乕偱摦偔僈儞僟儉偼丄偳傫側偵憗偔摦偄偰傕恖娫偺嬝擏偺傛偆側摦偒偼嵞尰偱偒側偄丅斀幩恄宱偺僗僺乕僪偵偼捛偄偮偗側偄偺偩偐傜丅
恖娫偺嬝擏偼乽廂弅乿偟偐偟側偄丅偙偺婡峔偵傛偭偰曏偺傛偆偵摦偔偺偩丅偩偐傜慺憗偄摦偒偑偱偒傞丅偁偭丄曏偲偄偆尵梩偱傂傜傔偄偨偧丅偝偭偒丄戞侾抜奒乣戞俁抜奒偱丄堦婥偵90搙傑偱偄偭偰丄僗僩僢僾偼偐偗傜傟側偄偲彂偄偰偼傒偨偗偳丄偦傟傕壜擻偐傕偟傟側偄丅側偤側傜丄怳傝壓傠偟偨曏傪巭傔傞偲偒偼庤庱偺僗僫僢僾傪棙偐偣偰丄斀懳曽岦偵僽儗乕僉傪偐偗傞丅偦偙偱乽價僢僔乿偲側傞傢偗偩丅嬝擏偱偙傟傪傗傞偨傔偵偼漢峈嬝偑兞偱摥偗偽椙偄偺偩偐傜丄戞堦偺愢傕傑偩桳岠偩偹丅偄偢傟偵偣傛係俆壄擭慜偵惗傑傟偨抧媴偑丄俁俆壄擭偐偗偰乽摦偔乿偙偲傪恑壔偝偣懕偗偨惉壥偑偙偙偵偁傞偺偩乮偍偍偘偝偐側乕乯丅曏偺傛偆側曏栄偺摦偒偑嬝擏傊偲恑壔偟偨偺偐側乕丅
偁偨偭偰偄傞偐偳偆偐偼抦傜側偄傛丅壗偟傠丄偙偙傑偱偔傞偲婎慴揑側夝朥妛傗惗棟妛偺杮偵偼壗傕彂偐傟偰側偄丅偲偄偆傛傝傕妛幰偝傫偨偪偼峫偊偨偙偲偁傞偺偐傕媈傢偟偄丅婽堜愭惗偼乽塣摦宯偺尋媶偑側偄乿偲扱偐傟偨偑丄俀俆擭屻偺崱傕偁傫傑傝曄傢偭偰側偄偺偐傕偟傟側偄丅偙傟傪杮奿揑偵挷傋傞偨傔偵偼惗暔妛偲忣曬岺妛偲暔棟岺妛偺壔妛僪僢僉儞僌偲偄偆偐丄嫟摨尋媶偑昁梫偩偹丅傕偆偳偭偐偱傗偭偰傞偩傠偆偗傟偳丅
弖娫揑側僗僺乕僪偺懍偄摦嶌偵僽儗乕僉傪偐偗傞曽朄偼傕偆堦偮偁傞丅偦偙偱侾倐側偺偩丅偙傟傕懌愳偝傫偵擟偣偪傖偄傑偟傚偆丅
丂壗偺偙偲偐夝傜側偄偲巚偄傑偡丅梫偡傞偵丄嬝偑廂弅偟偡偓側偄傛偆偵摨嬝偺兛嵶朎偵梷惂揑偵摥偔宱楬偑偁傞傢偗偱偡丅帺傜嬝傪廂弅偝偣傞傢偗偱偡偑丄偁傑傝偵傕嫮偄廂弅偼帺恎傪抐楐偟偐偹傑偣傫丅偦偙偱丄帺暘乮兛嵶朎乯偵僽儗乕僉傪偟偮偮帺暘乮兛嵶朎乯傪摥偐偣傞傢偗偱偡丅偆傑偔挷愡偟偰偄傞傢偗偱偡偹丅
丂偙偙偱搊応偡傞偺偑丄乬銯朼悗乭偲乬侾b慄堐乭偱偡丅嬝朼悗偵偼侾a慄堐偑拝偄偰偄傑偟偨偑丄銯朼悗偵偼侾b慄堐偑拝偄偰偄傑偡丅婎杮揑偵偼偙偺傆偨偮偼帡偰偄傞偺偱偡偑丄偪傚偭偲堘偆偲偙傠偑偁傝傑偡丅椉幰偲傕挘椡傪姶偠偨傜偦傟偧傟侾a丒侾b偑敪壩偡傞偺偱偡偑丄銯朼悗偼嬝偑怢挘偝傟偨偲偒傕廂弅偟偨偲偒傕摥偔偺偱偡丅嬝朼悗偼嬝偑怢挘偝傟偨偲偒偵偟偐摥偒傑偣傫偱偟偨偹乮悘堄揑嬝廂弅偵偍偄偰偼兞偵傛偭偰嬝朼悗偑廂弅偡傞偐傜侾a傕敪壩偱偒傞偺偱偡偑丒丒丒乯丅偱偼丄側偤嬝偑怢挘偝傟偨偲偒傕廂弅偟偨偲偒傕侾b偼敪壩偱偒傞偺偱偟傚偆偐丠丅偦傟偼丄銯朼悗偑嬝偵懳偟偰捈楍偵拝偄偰偄傞偐傜偩偲偝傟偰偄傑偡丅偦偆偡傟偽丄廂弅偵懳偟偰傕怢挘偵懳偟偰傕挘椡傪姶偠傞偙偲偑弌棃傞傢偗偱偡丅夝傜側偄偲巚偆偺偱丄壓恾丅
僀儊乕僕偱偒傑偡偐丠丂崪晅拝晹偲崪晅拝晹傪堷偒棧偡乮嬝偵偲偭偰偼怢挘偝傟傞乯偲偒偼嬝朼悗偵傕銯朼悗偵傕挘椡偑壛傢傞偱偟傚偆乮僄僉僗僷儞僟乕偺僶僱偺傛偆偵乯師偵丄嬝偑廂弅偟偨偲偒偼偳偆偱偟傚偆丠丂壓恾丅
嬝偑廂弅乮摨帪偵崪晅拝晹偼愙嬤乯偟偰傕銯朼悗偵偼挘椡偑壛傢傞偱偟傚偆乮嬝朼悗偼偨傞傓偗偳乯丅偙偆傗偭偰嬝偑廂弅偟偨偲偒傕丄怢挘偝傟偨偲偒傕銯朼悗偼挘椡傪姶偠偰侾b偼敪壩偱偒傞傢偗偱偡丅
侾b偼敪壩偟偨傜丄愐悜傑偱峴偭偰僯儏乕儘儞傪堦偮夘偟偰摨嬝偵偮側偑傞兛嵶朎偵怣崋傪憲傝傑偡丅偙偺娫偵嫴傫偩僯儏乕儘儞偼兛嵶朎偵梷惂揑偵摥偔傢偗偱偡丅愢柧偟朰傟傑偟偨偑丄銯朼悗偼傕偪傠傫銯偺拞偵懚嵼偟傑偡丅
兞塣摦慄堐丂乮嬝朼悗塣摦慄堐乯
崪奿嬝偺挿偝傪庴梕偡傞崪奿嬝偺姶搙傪挷愡偡傞恄宱慇堐丅偙偺慄堐傪墦怱惈偵憱傞怣崋偑丄嬝朼悗偵払偟丄嬝朼悗偺拞偵懚嵼偡傞廂弅惈偺峔惉惉暘傪廂弅偝偣傞偨傔偵丄嬝朼悗偺姶搙偑忋偑傞偲峫偊傜傟偰偄傞丅嬝朼悗偑嬝偺挿偝傪庴梕偡傞偵嵺偟偰丄挿偝偺帪娫揑曄壔乮怢挘懍搙乯偵懳偡傞姶搙偲丄挿偝偦偺傕偺偵懳偡傞姶搙偑栤戣偵側傞偑丄兞塣摦慄堐偵偼兞侾偲兞俀偺俀庬偑偁傝丄兞1偑怢挘懍搙偵丄兞2偑挿偝偺姶搙忋徃偵娭梌偡傞偲偄傢傟偰偄傞丅
兞宯
兞塣摦慄堐偺妶摦偼丄嬝挿庴梕婍乮嬝朼悗乯偺姶搙傪挷愡偡傞偙偲偵傛偭偰丄嬝廂弅偺嫮偝偵斀塮偡傞丅偟偨偑偭偰丄塣摦偵娭偟偰偼丄嬝偵僀儞僷儖僗傪憲傝崬傓兛塣摦僯儏乕儘儞偺傒偱側偔丄兞塣摦僯儏乕儘儞偺妶摦偑廳梫側堄媊傪帩偮丅兞塣摦僯儏乕儘儞傪嬱摦偡傞宯傪兞宯偲偄偆丅
兞亅挷愡
丂愐悜偺慜妏偵偼丄崪奿嬝傪巟攝偡傞塣摦僯儏乕儘儞乮兛僯儏乕儘儞乯偺傎偐偵丄傗傗彫宆偺僯儏乕儘儞乮兞僯儏乕儘儞乯偑偁偭偰丄偦傟偐傜弌偨恄宱慄堐乮俙兞慄堐乯偼兛僯儏乕儘儞偺恄宱慄堐乮俙兛慄堐乯偲堦弿偵憱偭偰嬝偵帄傞丅
丂兞僯儏乕儘儞偺栶栚偼嬝朼悗偺拞偺嬝慄堐乮悗撪嬝慄堐乯傪條乆側掱搙偵廂弅偝偣偰丄怢揥巋寖偵懳偡傞姶搙傪挷愡偡傞偙偲偵偁傞丅偙傟傪兞挷愡偲偄偆丅
丂怢揥巋寖偑庛偄偲偒偱傕丄悗撪嬝慄堐偑廂弅偟偨忬懺偱嶌梡偡傞偲丄偁偨偐傕怢揥巋寖偑嫮傑偭偨偐偺傛偆偵嬝朼悗偺姶妎恄宱乮侾倎孮慄堐乯偵僀儞僷儖僗偑憹壛偟丄偙傟偑兛僯儏乕儘儞偵揱偊傜傟傞偺偱怢挘斀幩偑嫮偔偍偙偭偰偔傞丅偙傟偼兞挷愡偺堦椺偱偁傞丅
丂忋埵擼偐傜偺壓峴惈巜椷偵傛偭偰嬝廂弅偺掱搙傪壛尭偟傛偆偲偡傞偲偒丄巜椷偼傑偢兞僯儏乕儘儞偵嶌梡偟丄擇師揑偵兛僯儏乕儘儞偺妶摦偑曄壔偡傞偲偄偆帠椺傕抦傜傟偰偄傞丅
兞娐乮兞儖乕僾乯
兞塣摦慄堐偺妶摦偼丄嬝挿庴梕婍乮嬝朼悗乯偺姶搙傪忋偘傞丅嬝朼悗偐傜惗偠偨怣崋偼丄拞悤恄宱宯偵岦偐偄丄塣摦僯儏乕儘儞偵擖椡傪梌偊傞丅塣摦僯儏乕儘儞偺弌椡偼丄嬝廂弅傪堷偒婲偙偟丄嬝朼悗偼嬝慄堐偺拞偵崿嵼偡傞偺偱丄椡妛揑偵塭嬁傪庴偗傞丅偙偺傛偆偵丄拞悤恄宱宯偲嬝偲偺娫偵偼媮怱惈宱楬偍傛傃墦怱惈宱楬偺儖乕僾偑偁傝丄偙偺宱楬傪捠傞怣崋偑丄嬝偺挘椡傪挷愡偡傞丅偙偺挷惍偵兞塣摦慄堐偺妶摦偑娭梌偟偰偄傞偺偱兞娐乮兞儖乕僾乯偲傛偽傟傞丅
兞峝捈亖兞屄弅
拞悤恄宱宯偵壗傜偐偺尨場偑偁偭偰丄兞塣摦僯儏乕儘儞乮兞宯乯偺擖椡偑憹戝偡傞偲丄崪奿嬝偺挿偝偺庴梕婍偱偁傞嬝朼悗偺姶搙偑崅傑傞丅偙偺偙偲偵傛傝丄崪奿嬝偵傢偢偐偺怢挘傪梌偊偨偲偒偵懡戝偺挘椡偑惗偢傞忬懺偲側傝丄娭愡偼偒傢傔偰摦偐偟偵偔偔側傞丅偙偺忬懺傪兞屄弅偲傛傃丄怢嬝宯偵惗偢傞偺偑捠忢偱偁傞丅乮堛妛戝帠揟乯
丂
尵梩偱棟夝偡傞傛傝恾傒傞曽偑憗偄
戞11恾
偙傟偑兞儖乕僾偲偄傢傟傞夞楬偩丅嬝擏偺塣摦偵偼忢偵偙偺兞偑愭峴偟偰婲偙偭偰偄傞丅
嵟弶偺巜椷乮戞侾抜奒乯偼擼偐傜兞嵶朎偵撏偒丄偦傟偑嬝朼悗傪廂弅偝偣丄廂弅偝傟偨嬝朼悗偼姶搙傪忋偘偰嬝擏偺忬懺傪抦傝丄嬞挘偑昁梫側傜偽侾倎偵怣崋傪棳偟丄偦傟偑兛嵶朎傪嫽暠偝偣丄嬝擏偺廂弅傪婲偙偝偣傞丅
嬝偑廂弅偡傞偲丄偦偺怣崋偼偙傫偳偼兞傪偼偢偟丄捈愙嬝朼悗偵払偡傞丅偦偙偐傜戞俀抜奒偺巒傑傝偲側偭偰弞娐偑婲偙傞丅
偦偺弞娐偼昁梫偑柍偔側傞傑偱壗搙傕懕偗傜傟傞偙偲偵側傞丅怴偨側昁梫偑惗傑傟傞偲戝擼偐傜兞傊偺巋寖偑嵞傃婲偙傝丄嬝擏傪怴偨側僗僥乕僕傊偲摫偄偰峴偔偙偲偵側傞丅偙傟偑丄慜偵僐儊儞僩乮庤媄椕朄偺尨棟乯偱偼愢柧偡傞偙偲偺偱偒側偐偭偨嬝擏偺僼傿乕僪僶僢僋儖乕僾偩丅
偙偺傛偆偵傒偰傒傞偲丄嬝朼悗偼僽儕僞僯僇昐壢帠揟偱弎傋傜傟偰偄傞扨側傞僗僩儗僀儞丒僎乕僕乮傂偢傒寁乯偺挘椡専弌慺巕偱偼側偔丄婲摦僗僀僢僠偱偁傝丄娔帇儌僯僞乕偱偁傝丄巌椷搩偺栶栚傕偟偰偄傞丅偝傜偵嫽枴怺偄偙偲偼丄偙偺兞嵶朎傊偺巜椷偼丄擼偺塣摦栰係丄俇丄彫擼丄戝擼婎掙妀丄擼姴栄條懱側偳偺庡梫側拞悤偐傜婲偙偭偰偄偄傞偙偲偱偁傞丅
戞12恾
偐偭偰庤媄椕朄偺嫄惎偱偁傞栰岥愭惗偲婽堜愭惗偼嬝擏偵堎忢側嬞挘傪婲偙偝偣偰偄傞尨場傪悕懱奜楬偺栤戣偲偟偰媮傔傛偆偲偟偨丅栰岥愭惗偼偦傟傪愽嵼堄幆偺栤戣偲偟偰偲傜偊丄婽堜愭惗偼斀幩偺堎忢偲偟偰偲傜偊傛偆偲偟偨丅
偦偺悕懱奜楬偺巋寖偼兞嵶朎傪揱偊傜傟偰偄傞偺偱偁傞丅愽嵼堄幆偺嵗傪擼偵嫮偄偰媮傔傟偽丄偦傟偼戝擼偺屆旂幙偲偄偆偙偲偵側傞偑丄偙偺兞傪巋寖偡傞擼偺嵗偵偼丄屆旂幙憌偐傜偺敪壩傕偁傞偐傕偟傟側偄丅傑偨丄僼傿乕僪僶僢僋偺堎忢偲偟偰媮傔傛偆偲偟偨婽堜愭惗偵偼兞儖乕僾偲偄偆夞摎偑梌偊傜傟傞偺偱偼側偐傠偆偐丅
埲忋偺偙偲偐傜孹幬埑偺惗傒弌偟偰偄傞偺偼丄偙偺兞偺夞楬側偺偩偲巚偆丅婽堜愭惗偼偙傟傪愢柧偟傛偆偲偟偰偄偨偺偱偼側偐傠偆偐丅偙偙傑偱撉傑傟偨傜丄傕偆堦搙丄栠偭偰婽堜愭惗偺嬝朼悗偺愢柧傪尒捈偟偰梸偟偄丅丒丒丒丒丒慡晹偼暘偐傜側偄偲偟偰傕丄壗偲側偔丄揱偊偨偐偭偨偙偲偺暤埻婥偼暘偐偭偰偒傑偣傫偐丅
亀偱偼昦婥傪恌傞偲偒偵丒丒丒娭愡偑堎忢傪婲偙偟偨傝丄摦偐側偔側偭偨傝偟偨偲偒偵偼丒丒丒嬝朼悗側偺偩丅偙偺尨棟偑傛偔棟夝偝傟偰偄傞偲娭愡摍偺摦偐側偔側偭偨傕偺偺挷惍偼旕忢偵傗傝堈偄丅偙偺尨棟偑暘偐傜側偄偺偱偼偄偔傜挷惍傪壛偊偰傕帯傜側偄丅乮婡娭帍26崋乽巋寖偺婎慴尨棟乿乯亁偳偆偱偡丅
偝偭偒丄帯椕傪庴偗偰偄偭偨曽偑丄惍宍奜壢偱尅堷偟偨傜偟傃傟偑弌偰偒偨偲慽偊偰偄偨丅崱傑偱偺偺峫嶡偐傜丄尅堷椕朄傪峫偊偰傒傞丅goopon偝傫偺棟孅偐傜峴偗偽丄偮傑傝侾倎傪梷惂偡傞偵偼侾倐偺尅堷傪嫮傔傟偽椙偄偲偄偆寢榑偵側傞偗傟偳丄偦偆偡傟偽嵟椙偺帯椕朄偼尅堷椕朄偲偄偆偙偲偵側傞丅偱傕丄偡偱偵弎傋偰偒偨偲偍傝丄侾倎偼侾倐偱偼梷惂偝傟側偄丅壗傜偐偺宍偱嬝朼悗偵巋寖傪梌偊偰丄兞傪捠偠偰丄偡偱偵偁傞僼傿乕僪僶僢僋偺儖乕僾偵曄壔偑婲偙偭偨偲偒偵偺傒曄壔偡傞偺偩丅
偟偐偟丄尅堷椕朄偱偼帪偲偟偰丄傗傝曽偑埆偄偲丄嬝朼悗偵巋寖偑峴偐側偄偽偐傝偑丄傛偗偄嬝朼悗傪嫽暠偝偣偰丄嬝擏傪傛偗偄偵嬞挘偝偣埆壔偡傞偙偲偵傕側傝偐偹側偄丅傢傟傢傟偑孹幬埑傪娚夝偡傞僞乕僎僢僩偵偡傋偒傕偺偼丄偙偺嬝朼悗偲兞儖乕僾側偺偩丅乽庤媄偺摑堦尨棟偦偺俀亅斀幩曇亅乿偺拞偱弎傋偨乽嶖岆偵傛偭偰惗偠傞拞棫揰偺僘儗乿偼偙偺兞儖乕僾偱婲偙偭偰偄傞偺偩偲巚偆丅偙偺栤戣偙偦丄婽堜愭惗偑斢擭丄捛媮偟傛偆偲偟偰壥偨偣側偐偭偨斶婅傪壥偨偡尞側偺偩丅
2001.5.15乮壩乯